
Fritsch's Quagga Photo (1864) taken in South Africa Has Been Rediscovered
By Branden Holmes and Jasper Hulshoff Pol. Published on 27 April 2024.
Artist Jasper Hulshoff Pol (website), with minor assistance from Branden Holmes, and confirmed by Dr. Peter Heywood, has rediscovered a stereophoto slide copy of a live Quagga photographed by Gustav Theodor Fritsch (1838-1927) in South Africa on 8 April 1864 (Fritsch, 1868).
Introduction
Africa is most often thought of as the continent that retains the largest proportion of its Late Pleistocene (c.126ka-11.7ka) megafauna (≥45kg), with the so-called "Big Five" (lion, leopard, rhinoceros, elephant, buffalo) only the tip of the biological iceberg. Alongside zebras, giraffes, Nile crocodiles, hartebeest, gnu (wildebeest), hippos, Greater kudus, Bongos, Great White sharks, Nile perch, Humpback whales, etc. etc. But palaeontological work over the last few decades has uncovered numerous large mammal (>5kg) extinctions, which are concentrated around the Late Pleistocene-Holocene border (13ka-6ka) (Faith, 2014). With the extinction rate increasing again after European colonisation, with the loss of taxa such as the Bluebuck (Hippotragus leucophaeus) (Hempel et al., 2024), the Cape warthog (Phacochoerus aethiopicus aethiopicus) (Grubb & d'Huart, 2010), the Bubal hartebeest (Alcelaphus buselaphus buselaphus) (IUCN SSC ASG, 2017) and the Senegalese giraffe (Giraffa camelopardalis senegalensis) (Petzold et al., 2020). One of the most famous of these latter disappearances is the Quagga (Equus quagga quagga), not least due to the long-term project (The Quagga Project) attempting to return its quasi-phenotype to southern African savannahs and grasslands (Parsons et al., 2007). It was last recorded in the wild in 1872 (Gippoliti et al., 2018), and the endling died in the Amsterdam Zoo on 12 August 1883 (Willoughby, 1966). It is one of relatively few extinct taxa for which 19th century photos of living individuals exist, and one of few to be the subject of multiple books (Barnaby, 1996; Green, 1996; Spreen, 2016; Heywood, 2022), and this latest photographic find emphasises the fact that research into the Quagga is very much ongoing.
The London Photos: All That Were Taken?
There are five surviving photos (see Appendix) of a Quagga (Equus quagga quagga) mare who resided in the London Zoo from 1851-1872, with her year of death seemingly coinciding with that of the last wild individual (Gippoliti et al., 2018). They were taken by Frank Haes (1864) and Frederick York (1870) (Heywood & Dietrich, 2021), but the last of them was only published more than a century later in 1991 (Huber, 1991, 1994). And even then, the details of the photographs have been made clearer by further research (Edwards, 1996; Fuller, 2013; Heywood & Dietrich, 2021). Today these five photos are now all freely available on the internet as they are in the public domain, including as part of the Wikipedia article on the subspecies. But it is sometimes stated that she was the only Quagga ever photographed alive (e.g. Huber, 1994; Wikipedia). However, it has been known by a handful of researchers that Gustav Theodor Fritsch (1838-1927), a medical doctor, took at least one (and possibly multiple) photos of a living specimen at the farm of Andrew Hudson Bain, 'Quaggafontein', on 8 April 1864, which was used to produce a photoxylograph (Fritsch, 1868; Barnaby, 1996; Heywood & Dietrich, 2021):

Above: a photoxylograph of a Quagga in South Africa. Reproduced from (Heywood & Dietrich, 2021), who reproduced it from (Fritsch, 1868).
The production of a photoxylograph involves the photochemical transfer of a photographic image onto a woodcut block prior to carving, and the fate of the original photograph/s is unknown (Heywood & Dietrich, 2021). The background has been noted as less photograph-like (Heywood & Dietrich, 2021), suggesting that multiple photographs may have been taken that were used as reference material for the stone wall and window shutters by the woodblock carver (Ibid., p. 8):
"Most probably, in our view, the wood engraving used to produce Figure 1 was reconstructed from other photographic images of these animals as the stone wall in the background with the window and shutters was drawn separately from other reference material and looks less ‘photographic’ than do the animals."
However, it must be noted that Jasper is of the opinion that the photoxylograph was produced from this stereophoto.
The Rediscovery of Fritsch's (1864) Photo
In December 2022, painter Jasper Hulshoff Pol noticed a listing for a slide of a Quagga in a July 1879 catalogue from Max Fritz (Fritz, 1879:8), and set about trying to track it down in case it was a 'new' photograph. Consequently, I sent him the Heywood & Dietrich (2021) paper about the photoxylograph, hoping that it might turn out to be a copy of the original photo. After reading the paper, Jasper searched for Gustav Fritsch and came across an eBay listing of a stereophoto of a zebra made by Gustav Fritsch. He purchased it, and when it arrived he noticed that it was the same image as Fritsch's photoxylograph, only mirrored due to the transfer process involved in photoxylography. I suggested that he contact Dr. Peter Heywood, who happily confirmed that the slide depicts the same Quagga as the photoxylograph (Peter Heywood, pers. comm. to Jasper Hulshoff Pol, December 2022):

Above: the front of the slide. Kindly provided by Jasper Hulshoff Pol.

Above: the reverse of the slide. Kindly provided by Jasper Hulshoff Pol.
A Stereophoto: One Photo, Two Images
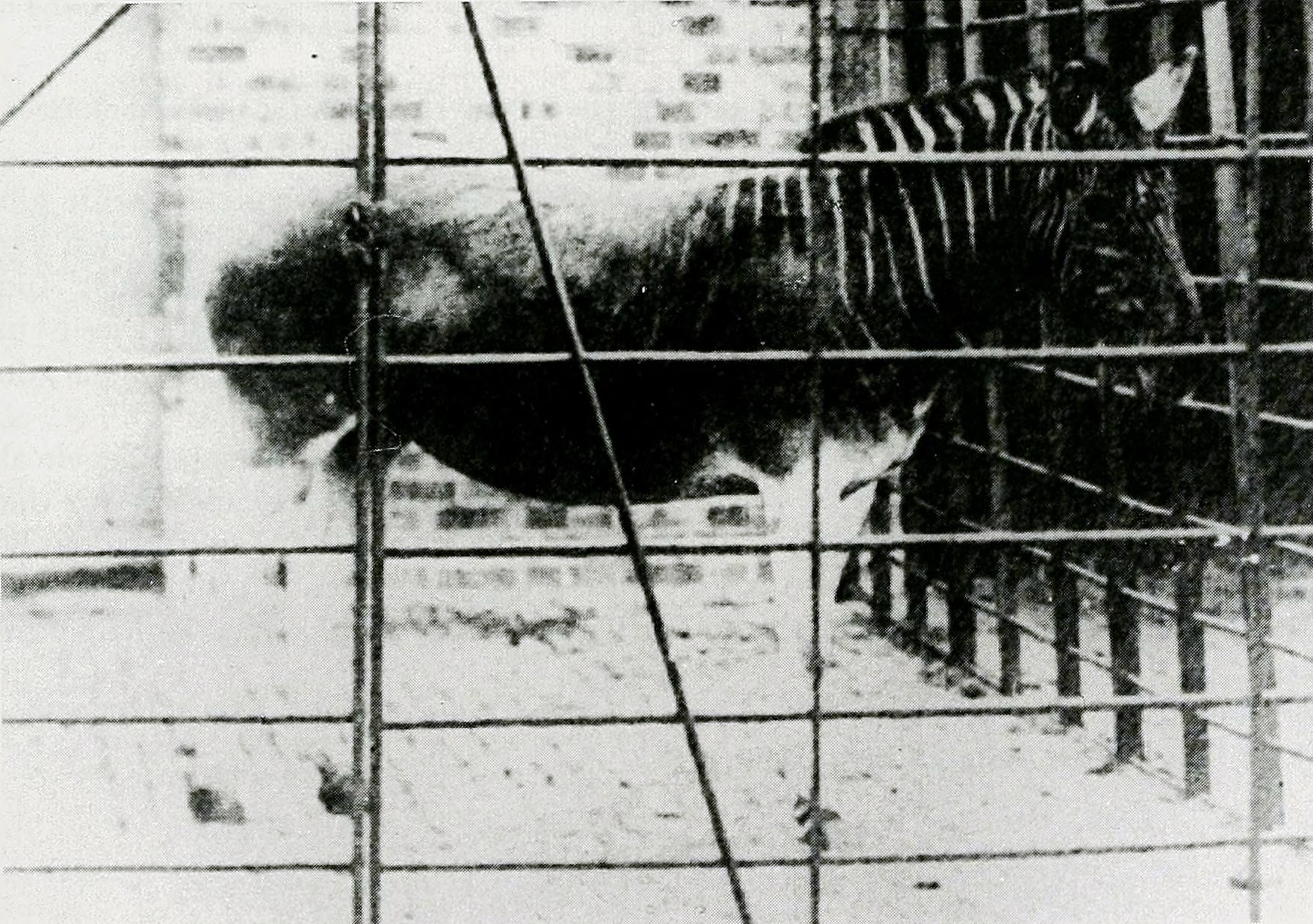
The original Max Fritz catalogue listing (Fritz, 1879:8) appears to be for a magic lantern slide for use in projection, which Jasper's slide copy is not. The original eBay listing for Jasper's slide stated that it was a stereophoto, implying that it was taken with a camera that uses two lenses centimetres apart to simulate our slightly different left and right eye positions (see Sleightholme et al., 2016:Fig. 4). When a stereophoto is viewed through a stereoscope it produces a three-dimensional image (Ibid.). This is consistent with some minor differences between the two images that can be seen below, showing that they are taken from slightly different angles: 1) The front legs are more open in the right image, 2) the white object on the ground between the rear legs is obscured more in the left image, 3) the shadow on the back of the right front leg is a different shape, and 4) the duck under the Quagga is in a different position relative to the front and back legs of the latter. Several other possible differences are harder to confidently distinguish given the blurrier quality of the left image.

Above: a side-by-side comparison of the two frames of the slide, with minor differences between them noticeable. Indicative of a stereophoto, in line with the original eBay listing.
Fritsch' Zebra, A True Quagga?
Heywood & Dietrich (2021) provide two lines of evidence to argue that the zebra Fritsch photographed on 8 April 1864 was a true Quagga and not a 'bonte' (striped) Quagga (i.e. Burchell's zebra, E. q. burchelli). The first was the degree of lack of stripes that goes beyond known cases of lesser-striped Burchell's zebra (i.e. no leg striping and reduced body striping), and the second was Fritsch's precision of language in describing the animal as a Quagga and not a bonte Quagga in full knowledge of the difference (Ibid.). Upon receiving a copy of the slide from Jasper via email, Dr. Heywood raised the fact that the slide lists the animal as a 'bonte Quagga' (i.e. well striped Burchell's zebra) and disagreed with this designation, reiterating the first line of argumentation made in his joint paper:
"Clearly, it's the same quagga illustrated in our paper and yet the description terms it "bonte" (striped) and names it as Equus burchelli. I have to disagree with this designation as leg stripes are absent and the body striping is reduced. In fact, the degree of body striping is similar to that in several museum specimens and illustrations labelled as "quagga.""
(Peter Heywood, pers. comm. to Jasper Hulshoff Pol, December 2022)
Conclusion
The evidence shows that Gustav Theodor Fritsch took a stereophoto (that produced two images from slightly different angles) of a Quagga on 8 April 1864 in South Africa. With Jasper's discovery of two images on a single stereoscopic slide bringing the total number of images of living Quaggas to at least eight or none: five photographs from London including at least one stereoview (resulting in at least six images), and one stereophoto (two images) and one photoxylograph (one image) (for nine total images), or, one stereophoto (two images) that were used to produce a photoxylographic version of them (for a total of eight images).
The fact that the reverse of the slide falsely identifies the zebra as a 'bonte Quagga' (Equus burchelli) may have prevented the significance of the slide from being recognised earlier. Although copies of such slides must now be relatively rare and not likely to be encountered very frequently. This raises the important possibility that other visual depictions of living Quaggas may await discovery/recognition, so long as the degree of lack of striping can be sufficienty seen to rule out lesser-striped examples of Burchell's zebra (Equus quagga burchelli).
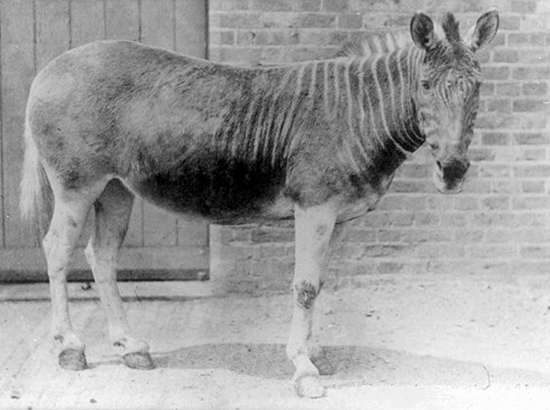
Appendix: The five London photographs (1863-1870)


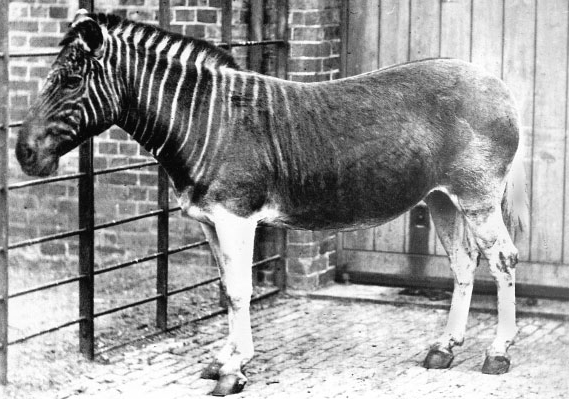
Above: one of two images from the stereoview (see here).
References:
Barnaby, David. (1996). Quaggas and Other Zebras. Plymouth: Bassett Publications. 114 pp.
Edwards, John. (1996). London Zoo from Old Photographs: 1852-1914. London: John Edwards. 240 pp.
Faith, J. Tyler. (2014). Late Pleistocene and Holocene mammal extinctions on continental Africa. Earth-Science Reviews 128: 105-121.
Fritsch, Gustav Theodor. (1868). Drei Jahre in Süd-Afrika. Reiseskizzen Nach Notizen des Tagebuchs zusammengestellt. Breslau (Ferdinand Hirt). 416 pp.
Fritz, Max. (1879). Das Scioptikon vervollkommneter Projectionsapparat für den Unterricht: Fünfte Auflage (fifth edition): Juli (July) 1879. Goerlitz, Silesia: Max Fritz.
Fuller, Errol. (2013). Lost Animals: Extinction and the Photographic Record. London: Bloomsbury Press. 256 pp.
Gippoliti, Spartaco, Cotterill, Fenton P. D., Zinner, Dietmar and Groves, Colin P. (2018). Impacts of taxonomic inertia for the conservation of African ungulate diversity:an overview. Biological Reviews 93(1): 115-130. https://doi.org/10.1111/brv.12335
Green, Tamara. (1996). The Quagga (The Extinct Species Collection). Gareth Stevens Pub. 24 pp.
Grubb, Peter and d'Huart, Jean-Pierre. (2010). Rediscovery of the Cape Warthog Phacochoerus aethiopicus: a review. Journal of East African Natural History 99(2): 77-102. https://doi.org/10.2982/028.099.0204
Hempel, Elisabeth, Faith, J. Tyler, Preick, Michaela et al. (2024). Colonial-driven extinction of the blue antelope despite genomic adaptation to low population size. Current Biology. https://doi.org/10.1016/j.cub.2024.03.051
Heywood, Peter. (2022). The Life, Extinction, and Rebreeding of Quagga Zebras: Significance for Conservation. Cambridge University Press. 242 pp.
Heywood, Peter and Dietrich, K. H. (2021). A quagga photographed in Africa (Mammalia, Perissodactyla, Equidae). Spixiana 44(2): 209-211.
Huber, Walter. (1991). Ein bisher unveröffentlichtes Foto eines lebenden Equus quagga quagga Gmelin, 1788 (Mammalia, Perissodactyla, Equidae). Spixiana 14(2): 235-236.
Huber, Walter. (1994). Dokumentation der fünf bekannten Lebendaufnahmen vom Quagga, Equus quagga quagga Gmelin, 1788 (Mammalia, Perissodactyla, Equidae). Spixiana 17(2): 193-199.
IUCN SSC Antelope Specialist Group. (2017). Alcelaphus buselaphus ssp. buselaphus. The IUCN Red List of Threatened Species 2017: e.T813A50181474. https://dx.doi.org/10.2305/IUCN.UK.2017-2.RLTS.T813A50181474.en. Accessed on 04 July 2022.
Parsons, Rochelle, Aldous-Mycock, Colleen and Perrin, Michael R. (2007). A genetic index for stripe-pattern reduction in the zebra: the quagga project. South African J. of Wildlife Research 37(2): 105-116. https://doi.org/10.3957/0379-4369-37.2.105
Petzold, Alice, Anne-Sophie Magnant, David Edderai, Bertrand Chardonnet, Jacques Rigoulet, Michel Saint-Jalme & Alexandre Hassanin. (2020). First insights into past biodiversity of giraffes based on mitochondrial sequences from museum specimens. European Journal of Taxonomy 703: 1-33.
Sleightholme, Stephen R., Campbell, Cameron R. and Kitchener, Andrew C. (2016). Frank Haes' thylacine. Australian Zoologist 38(2): 203-211. https://doi.org/10.7882/AZ.2016.022
Spreen, Reiner. (2016). Monument voor de quagga. Uitgeverij Fusilli. [in Dutch]
Willoughby, D. P. (1966). The vanished quagga. Natural History 75(2): 60-63.
Updates to My Books
1. What's Lost and What Remains: The Sixth Extinction in 100 Accounts (2021, eBook)
4. The Frosted phoenix hasn't been seen since 1959 (Titanomis sisyrota)
Titanomis has been rediscovered on Stewart Island, where it was unknowingly photographed by Pav Johnsson on 2 March 2024, before being identified as Titanomis by Dr. Robert Hoare (who gave the species its vernacular name) as the long lost moth (Mitchell, 2024).
References:
Mitchell, Charlie. (2024, March 30). Mysterious moth unseen for 65 years accidentally rediscovered by tourist. The Press (Te Matatika). Available at: https://www.thepress.co.nz/nz-news/350225711/mysterious-moth-unseen-65-years-accidentally-rediscovered-tourist [Accessed 30 March 2024]
25. Anthropogenic extinction sucks (Xerobdella lecomtei)
The European land leech (Xerobdella lecomtei) is not in fact endemic to the birch forests around Graz, Austria, and is not thought to be extinct. I failed to pick up on the fact that Kutschera et al. (2007) used the term "local extinction", as it has also been reported from Slovenia (e.g. Jueg, 2015) & Italy (e.g. Nesemann & Neubert, 1999).
References:
Jueg, Uwe. (2015). Xerobdella praealpina Minelli, 1971 (Hirudinea, Xerobdellidae) in Österreich und Slowenien. Lauterbornia. 79: 145-149.
Kutschera, U., Pfeiffer, I. and Ebermann, E. (2007). The European land leech: biology and DNA-based taxonomy of a rare species that is threatened by climate warming. Naturwissenschaften 94: 967-974.
Nesemann, H. and Neubert, E. (1999). Branchiobdellida, Acanthobdellea, Hirudinea.- In: Schwoerbel, J. & P. Zwick (eds): Süßwasserfauna von Mitteleuropa. Begründet von A. Brauer 6(2), 178 pp., Heidelberg.
37. The man who cried shark? (Carcharhinus hemiodon)
The most recent RedList assessment for the species revises its last record back to 1960, with persistent issues relating to correct identification of individuals due to possible confusion with a number of other species (Kyne et al., 2022). The IUCN therefore does not recognise the alleged rediscovery of the species made by Jessica Evans, Forrest Galante's wife. Moreover, trouble with field identification makes it difficult to assign the species to a specific conservation category, emphasising that its status as missing is tentative at best. Thus again making it a poor choice for Galante to search for, as opposed to other species whose identification in the field is not problematic, including the greater part of biodiversity (invertebrates, plants and fungi) that he almost completes neglects.
References:
Kyne, P. M., Jabado, R. W., Akhilesh, K. V., Bineesh, K. K., Booth, H., Dulvy, N. K., Ebert, D.A., Fernando, D., Khan, M., Tanna, A. and Finucci, B. (2021). Carcharhinus hemiodon (errata version published in 2022). The IUCN Red List of Threatened Species 2021: e.T39369A221513674. https://dx.doi.org/10.2305/IUCN.UK.2021-1.RLTS.T39369A221513674.en. Accessed on 17 December 2022.
41. When native species must go (Lampetra minima)
At the time of writing, I was only aware of a single species of lamprey that is considered possibly extinct, a migratory species from Ukraine last recorded in the late 1800s (Eudontomyzon sp. nov. 'Migratory'). A recent IUCN RedList assessment has treated the Chapala lamprey (Tetrapleurodon spadiceus) as "Critically Endangered (Possibly Extinct)", which was endemic to Lago de Chapala and a connecting portion of the Rio Grande de Santiago, and last recorded in 1987 (Snoeks et al., 2019).
References:
Snoeks, J., Lalèyè, P. and Contreras MacBeath, T. (2019). Tetrapleurodon spadiceus. The IUCN Red List of Threatened Species 2019: e.T169396A132675777. https://dx.doi.org/10.2305/IUCN.UK.2019-2.RLTS.T169396A132675777.en. Accessed on 17 December 2022.
48. A giant in the canopy (Ctenomorpha gargantua)
In the book I wrote the following about Coxen's fig parrot (Cyclopsitta coxeni):
"It has been the subject of numerous unconfirmed sightings over the last few decades, although verified records do apparently exist (BirdLife International, 2021)".
But a recent paper reports that there have not been any independently verified sightings of the species since at least 2002 (Garnett et al., 2022).
References:
BirdLife International. (2021). Species factsheet: Cyclopsitta coxeni. Downloaded from http://www.birdlife.org on 15/08/2021.
Garnett, Stephen T., Hayward-Brown, Brittany K. et al. (2022). Australia's most imperilled vertebrates. Biological Conservation 270: 109561. https://doi.org/10.1016/j.biocon.2022.109561
52. An extinct sheep-sized echidna (Zaglossus hacketti)
Since the book's publication in 2021, a new paper (Flannery et al., 2022) has erected the new genus Murrayglossus for Zaglossus hacketti (now Murrayglossus hacketti) as well as engaged in some grounded speculation over its ecology:
"The ecology of M. hacketti is unknown. Yet it is morphologically distinct from the sympatric Tachyglossus and Megalibgwilia, implying some degree of niche partitioning. Murray (1978b, p. 53) commented that ‘one possible postural correlate of the shortened tibia and relatively long femur [in M. hacketti] is an adaptation to shift the centre of gravity of body mass backwards. This may have allowed mobility of the forelimbs for digging or tearing and may have permitted the animal to easily assume an assisted bipedal stance while feeding on ants or termites nests, a posture sometimes used by both living genera’. We would add that a semi-vertical posture might have also facilitated arboreality in M. hacketti. Surprisingly, Zaglossus bartoni is capable of ascending vertical fences (Flannery 1998). Furthermore, the vicinity of Mammoth Cave is densely forested. Arboreal-nesting termites (Calaby & Gay 1959) could have feasibly provided a viable food source. If correct, then niche separation among tachyglossids potentially involved Tachyglossus as a terrestrial feeder on colonial invertebrates, Megalibgwilia as a terrestrial feeder on large soil invertebrates (e.g., scarab beetle larvae: see Murray 1978b), and M. hacketti as a scansorial feeder on arboreal, colonial invertebrates." (Flannery et al., 2022)
References:
Flannery, Timothy F., Rich, Thomas H., Vickers-Rich, Patricia, Ziegler, Tim, Veatch, Grace and Helgen, Kristofer M. (2022). A review of monotreme (Monotremata) evolution. Alcheringa: An Australasian Journal of Palaeontology 46(1): 3-20. https://doi.org/10.1080/03115518.2022.2025900
64. When to declare? Cricketers and conservationists share a problem (Rhachistia aldabrae)
Since writing the book, a new preprint (viz. not peer-reviewed) has appeared which calls into question the claim that climate change has been responsible for the sharp decline in Rhachistia aldabrae (Altaba, 2022).
References:
Altaba, C.R. (2022). Extinction through Climate Change: Review of Evidence and Analysis of Two Land Snails from the Seychelles Islands. Preprints 2022, 2022100315 (doi: 10.20944/preprints202210.0315.v2).
86. Snaking its way out of extinction (Aipysurus foliosquama)
Wellington's solitary coral (Rhizopsammia wellingtoni) had not been recorded since 2000 (Hickman et al., 2007) when I wrote the book. Happyily, it was rediscovered in January 2024 (Keith et al., 2025).
References:
Hickman, C., Edgar, G. and Chiriboga, A. (2007). Rhizopsammia wellingtoni. The IUCN Red List of Threatened Species 2007: e.T63579A12683468. https://dx.doi.org/10.2305/IUCN.UK.2007.RLTS.T63579A12683468.en. Downloaded on 25 December 2015.
Keith, Inti, Gosliner, Terry and Albright, Rebecca. (2025). Rediscovery of Rhizopsammia wellingtoni in Galápagos after two decades. Marine Biology 172: 122. https://doi.org/10.1007/s00227-025-04679-4
88. Extinction is a burning problem (Phalanger matanim)
An online story claims that Michael Smith has found a family eating a Telefomin cuscus (Phalanger matanim) in presumably 2022 or 2023 (Sweetnam, 2023), which has reportedly been confirmed by Dr. Tim Flannery (Glen, 2023).
References:
Glen, Allan. (2023, March 4). British tourist spots three species thought to have been extinct. The Telegraph (online), available at: https://www.telegraph.co.uk/world-news/2023/03/04/british-tourist-holiday-papua-new-guinea-spots-three-species/ [Accessed 19 March 2023]
Sweetnam, James. (2023, March 4). Brit tourist baffles expert by discovering three species thought to be extinct. Daily Star (online), available at: https://www.dailystar.co.uk/news/latest-news/brit-tourist-baffles-expert-discovering-29372313 [Accessed 5 March 2023]
96. A not so egg-cellent identification (Genyornis newtoni)
A new paper has resolved the identity of the mystery layer of the eggs, and shown that it was in fact Genyornis after all (Demarchi et al., 2022).
References:
Demarchi, Beatrice et al. (2022). Ancient proteins resolve controversy over the identity of Genyornis eggshell. PNAS Preprint e2109326119. https://doi.org/10.1073/pnas.2109326119
2. Thylacine: The History, Ecology and Loss of the Tasmanian Tiger (2023, CSIRO Publishing)
One niggle arises over the wording of a chapter of mine ("Never far apart: picturing Paris's pair of pouched predators"). I wanted an alliterative title, and originally had 'prehistoric' in place of 'pouched', but I felt that that did not do justice to the species, especially in the face of "placental chauvinism". However, I fear that I replaced it with an equally unsuitable word ("pouched") which might suggest that both the female and male had genuine pouches. In reality, the male's "pouch" is nothing of the sort and intended to be able to retract the scrotum into for protection.
The Damage Forrest Galante Has Done to Conservation Biology
By Branden Holmes. Published 16 August 2020. Rewritten 14 October 2023.
Introduction
Forrest Galante grew up in Zimbabwe, and is the grandson of Gerald Summerfield whom Galante claims helped prove that Coelacanths aren't extinct (Provost, 2016). However, a quick Google search does not bring up any results that indicate this is true. Rather, the discovery of the first known living specimen encountered by humans is well documented and omits the mention of any such person. It was caught off East London (South Africa) aboard the trawler of Captain Hendrick Goosen in December 1938, bought by a provincial museum curator, Marjorie Courtenay-Latimer, and recognised as the long extinct coelacanth by eminent ichthyologist J. L. B. Smith (Smith, 1956; Weinberg, 2000). Judging by Galante’s comments, it’s clear that his grandfather played a role in securing one of the later specimens caught in the Comoros islands (Provost, 2016), which is still a significant association with the species.
Indeed, it seems that this connection was prophetic, as if he was earmarked for discoveries, firsts and records1. According to the About page [archived version] on his website:
"At age 14, Galante was the youngest person to ever lead an international canoe safari down the Zambezi River."
And with his rise to public notice since moving to the US, first as a television contestant on 'Naked and Afraid', he is now best known as the host of the two seasons of ‘Extinct or Alive’ on Animal Planet. According to the Discoveries page on his website, the show resulted in Galante and his team rediscovering no less than eight species previously thought to be extinct, which would arguably make him the most prolific rediscoverer of species in the history of conservation biology. However, there is no evidence that he has actually rediscovered even a single species. Instead, he has a track record of taking credit for the work of others, ignoring recent records of species that he claims are missing, has never published any of his alleged rediscoveries in a peer-reviewed paper, and does not seem to care about the fate of the species he has allegedly rediscovered (i.e. does not give any updates on their status at all, saying if anything that he has "passed on" the rediscoveries to local researchers).
There are certainly instances in which a single individual has genuinely made multiple rediscoveries, such as the botanist Dr. Brian du Preez (Wild, 2019; Swingler, 2023), and the malacologist Dinarte Teixeira (PhD student) (e.g. Cameron et al., 2021; D. Teixeira pers. comm., 19 May 2022). But the successes of these individuals, as well as the rediscoveries made by the many other (citizen) scientists, are yet dwarved by those innumerable failed efforts to search for (possibly) extinct taxa that did not result in rediscovery. Not to mention the fact that most biologists never rediscover even a single taxon despite years of trying (whether explicitly or opportunistically). The fact that Galante allegedly rediscovers species so frequently, and in such short search windows, should immediately raise serious doubts over their veracity. And raise questions as to whether the false rediscovery narratives he constructs and pushes are negatively affecting the public's perception of the role, and competence, of conservationists more broadly, including those in poor and developing countries.
Red Flags: A Mere Media Personality?
Forrest Galante’s first public foray into putatively extinct species was not a published piece of research, but as a co-host of a one-off documentary aired in 2016 in search of the thylacine or Tasmanian tiger (Thylacinus cynocephalus) in Tasmania, called 'Extinct or Alive: Tasmanian Tiger'. It was evidently the inspiration for the title of his most well known television project, ‘Extinct or Alive’ (2018-2021). Mike Williams2 reminds me that one scene from the documentary has Galante interviewing a descendent of a man who shot a thylacine in the 1950’s and shows Galante a photo as proof. But this photo is actually one of a series taken in May 1930 depicting the thylacine shot by Wilf Batty on his family’s property at Mawbanna in Tasmania's north-west, the last known wild killed specimen. The fact that Galante does not pick up on this is disconcerting as it comprises one of the most famous series of photos of one of the most famous recently extinct species, and one which he evidently has a special interest in.
Forrest's website states that he has a bachelor's degree in biology from UC Santa Barbara with "special emphasis in marine biology and herpetology". Yet despite his education he makes numerous basic errors in his three appearances on The Joe Rogan Experience (JRE) podcast (as shown below), as well as in other media appearances including his own content, in line with the fact that he appears to (predominantly) earn a living as a media personality and not as a professional biologist, and certainly not someone who earns a living from researching and publishing (there are no peer-reviewed papers to his name). A biology degree is a commonly studied degree given the importance of the natural world, but it does not make one an expert biologist, as it broadly teaches you what we know about living organisms. Expertise requires further, active research on the part of the researcher over a number of years. For professional biologists, a standard part of which is conducting studies (whether in the field, using computer simulations, studying museum specimens, molecular work etc.) and getting manuscripts through the peer-review process to have their findings published.
Someone who is not a professional biologist can of course still make a significant contribution to biology, and the practical nature of the subject means its history is stuffed with self-taught individuals who have made major contributions. The rediscovery of a species is a clear example. So someone like Galante who has a bachelor's degree in biology is at a distinct advantage over those who never even attended university/college, and thus is very well placed to be such a contributor. Yet, far from displaying a deep level of knowledge regarding his speciality (large animals), he consistently makes basic errors. By way of just a few of the many examples from his various appearances on Joe Rogan's podcast, he states that the leopard (Panthera pardus) is larger than the jaguar (Panthera onca) (episode #1240, roughly 40:00 into the conversation). In reality the relationship is precisely the opposite, with the Jaguar being the third largest big cat in the world (Rabinowitz, 2005). He also strongly implies that since tortoises cannot swim large distances, inter-island travel is impossible (episode #1403, 1:05:50 into the conversation). In reality, it is well known that giant tortoises float (Patterson, 1973), and can even be found hundreds of kilometres from their home (e.g. Gerlach et al., 2006). And while their travel direction is thus contingent upon currents, the potential for inter-island travel is certainly there. But Forrest's third appearance on the podcast contains his biggest factual errors to date. For example, at the timestamp 1:42:06 in following video:
Forrest clearly claims that 1) the facial tumour disease in question is shared by "a lot of the animals" in Tasmania and thus is not limited to the Tasmanian devil (Sarcophilus harrisii), 2) it is herpes, and 3) it is due to an overabundance of prey, at least in the case of the Tasmanian devil. In reality, the disease is a transmissible cancer called Devil Facial Tumour Disease (DFTD) and first arose via a tumour in a female devil between 1977 and 1987 in north-east Tasmania (Patton et al., 2020; Jones, 2023 and references therein; Woods, 2023 and references therein). It is certainly not herpes, it has not been reported in any other native Tasmanian animal, and it is not due to the absence of the thylacine or overabundance of prey or both. It is transferred through the species' tendency to bite each other on the face, or rather both strains of DFTD, since a second strain (DFTD2) sadly arose in 2014 (Jones, 2023; Woods, 2023).
He also claims that the Reticulated python (Broghammerus reticulatus) is the heaviest snake in the world, while the Anaconda (presumably, the Green anaconda (Eunectes murinus)) is the longest (starts at 6:58 in the below video).
Reliable measurements for both species are scarce in comparison to exaggerated and unverified claims, but it is generally accepted that the Reticulated python (Broghammerus reticulatus) is the longest snake, while the more massive Green anaconda (Eunectes murinus) is significantly heavier. The longest reliably measured wild Reticulated python was 6.95m long, and weighed 59kg after not eating for an estimated 3 months (Fredriksson, 2005). While the longest of 780 wild Green anacondas was 5.21m, and the heaviest was 97.5kg (Rivas, 2000:Table 3-1).
Extinct or Alive? Episodes in the Forrest Galante Saga
Even given the numerous basic errors Galante makes, if he rediscovered even one species his name would be etched into the annals of conservation biology for all time. The fact that he claims to have been a part of no less than eight of them, documented during the two seasons (18 episodes) and five specials of the show 'Extinct or Alive', means we need to unbiasedly investigate their veracity, in case he possesses unique knowledge on how to find putatively extinct species that can be applied more broadly. But multiple issues arise immediately.
Non-existent targets
The list of 22 target (sub)species contains four whose taxonomic validity is seriously disputed: the Zanzibar leopard (Panthera pardus adersi) (Pakenham, 1984; Miththapala et al., 1996; Uphyrkina et al., 2001; Sun et al., 2023 [preprint]), the Newfoundland wolf (Canis lupus beothucus) (Nowak, 1995), the Formosan clouded leopard (Neofelis nebulosa brachyurus) (Buckley-Beason et al., 2006; Kitchener et al., 2006, 2017), and the Southern Rocky Mountain wolf (Canis lupus youngi) (Nowak, 1995). That is, they probably never existed as distinct forms in the first place. And you cannot rediscover what you cannot discover: a non-existent (sub)species. These episodes could have been better served looking for real species, especially given the opportunity to highlight the plight of smaller animals, particularly invertebrates.
Bias towards larger species
The list of target species (n=22) were without exception large-bodied vertebrates, with not a single invertebrate in sight. Perhaps if there were a reason to suppose that such large-bodied species deserve special emphasis, such as by having suffered catastrophic rates of recent extinction compared with smaller-bodied animals, then this bias might be justified. But such a bias is not sustained by the evidence, as the database on this website shows. Large-scale funding to search for putatively extinct species may have only been made available to target large, charismatic species, but there are plenty of invertebrates in his home state of California that could have been the subject of an episode for very cheap. For example, the Sacramento Valley hairy-necked beetle (Cicindela hirticollis abrupta) last seen around 1984, a pair of weevils last recorded in 1975 (Trigonoscuta rossi and T. yorbalindae), the Antioch robberfly (Cophura hurdi) from the unique Antioch dunes, or the Killerdana trapdoor spider (Aptostichus killerdana) last recorded on 15 February 1971.
Species not even missing, or already rediscovered
At the time of filming the respective episodes, three of the target species were not even missing: Pondicherry shark (Carcharhinus hemiodon), Whitetip weasel shark (Paragaleus leucolomatus), and Flapnose houndshark (Scylliogaleus quecketti). While three others had already been rediscovered by other people: Miller's grizzled langur (Presbytis canicrus), Cape lion (Panthera leo melanochaitus), Rio Apaporis caiman (Caiman crocodilus apaporiensis). Thus, six of the 22 target species had recent records, making it far, far more likely that he would succeed in "rediscovering" these not-actually-extinct targets that he claims to be possibly extinct. It is thus no surprise that all six of these feature among the list of eight targets that he has (helped) rediscover as part of the show. Only two (sub)species might truly have been rediscovered as part of the show: the Zanzibar leopard (Panthera pardus adersi) and the Fernandina Island giant tortoise (Chelonoidis phantasticus). But the former likely does not exist (see 'Non-existent targets' above), and the latter was rediscovered by another indiviual (Jeffreys Málaga) which Galante tried to steal credit for.
Season 1, Episode 1: Zanzibar leopard (Panthera pardus adersi)
The Zanzibar leopard (Panthera pardus adersi) was described by Reginald Pocock in 1932 (Pocock, 1932), and was endemic to the small and populous island of Unguja (1,666 km2 or 643 sq mi) in the Zanzibar archipelago off East Africa. However, the species was officially persecuted for it’s alleged links to witchcraft (Walsh & Goldman, 2012), with the last confirmed records seemingly dating to the 1980’s. However, the validity of the subspecies has also long been disputed (Pakenham, 1984; Miththapala et al., 1996; Uphyrkina et al., 2001; Sun et al., 2023 [preprint]), and was subsumed into the nominate subspecies (P. p. pardus) by (Miththapala et al., 1996). Given the serious questions over the validity of the subspecies, it was not a good choice to target.
Nevertheless, according to the Discoveries page on his website:
“In 2018, Galante captured trail camera footage of a Zanzibar leopard, a big cat that had been classified as extinct for over 25 years— with support from Dr. Martin Walsh and Dr. Helle V. Goldman. During that same expedition, Galante also captured the first known video footage of a servaline genet in the region. ”
A blog post by Dr. Helle Goldman and Dr. Martin Walsh, the foremost researchers of the Zanzibar leopard over the last couple of decades, however, tells a very different story (Goldman & Walsh, 2018). I apologise for quoting almost the entire blog post, however it is important to do so as the full context needs to be understood here:
“The quest for the Zanzibar leopard is the running theme of the premiere of the Animal Planet infotainment series, “Extinct or Alive", which airs in the US on Sunday, 10 June. We participated in the production of this extended episode by providing background material about leopards and other wildlife on the island, as well as the island’s history and culture. Travelling to Zanzibar a week in advance of the film crew, we scouted filming locations, lined up interviews, liaised with local forestry staff, and obtained access to recent photographs of alleged leopard pugmarks and leopard kills (eviscerated goats).
In addition, we deployed 10 camera traps in various locations, getting excellent video footage of some of Zanzibar’s small carnivores in the wild: the Zanzibar servaline genet (Genetta servalina archeri), the African palm civet (Nandinia binotata), and the Zanzibar bushy-tailed mongoose (Bdeogale crassicauda tenuis). To our knowledge, none of these has been filmed on Unguja island before, so this was exciting.
…
They did, however, use footage of an interview with us and many clips of the other animals that we had camera-trapped, including superb daytime (and therefore colour) video shots of a servaline genet.
...
In the absence of verified evidence, like most authorities we now presume the Zanzibar leopard to have been extirpated, despite tantalising reports and claims to the contrary. But this shouldn't stop people from enjoying Animal Planet's expertly crafted and highly entertaining narrative.”
As acknowledged to some degree by Galante, Dr. Goldman and Dr. Walsh were instrumental in the success of the episode, but he does not elaborate on what this involved exactly. According to Dr. Goldman and Dr. Walsh, it was they who captured the Zanzibar servaline genet (Genetta servalina archeri) on film. Given the totality of evidence cited in this article and that of (Wight, 2020), I personally have no doubt that Dr. Goldman and Dr. Walsh are telling the truth.
The fact that Goldman and Walsh have never recorded the Zanzibar leopard despite visiting the small island many times over the years, including 10 camera traps deployed in their latest attempt, while Galante and crew managed to capture a leopard in the short time they were on the island, should raise suspicions that it is a feral animal or the footage was not in fact taken on Unguja. In terms of the footage itself, keeping in mind that the Zanzibar leopard is not a distinct subspecies, Dr. Goldman and others are sceptical of its authenticity as representing a Zanzibar leopard on Zanzibar (Unguja):
"In 2018 a crew of Animal Planet, during the recordings of a television program, obtained a camera trap video of an alleged Zanzibar leopard (Li 2018). Some authorities responsible for the Zanzibar leopard do not consider this film to be reliable evidence (H. Goldman pers. comm.; Goldman and Walsh 2018) and its diffusion on the internet has been restricted to the American television newsmagazine Inside Edition and some blogs devoted to paranormal and other alleged mysteries. On the other hand, the author has defended the authenticity of the film (F. Galante, pers. comm.). The video undoubtedly shows a leopard, but the images do not allow to verify precisely the pattern of the rosettes or to determine the shooting’s locality. It should also be noted that a feral African leopard released in Zanzibar is an option which can be eliminated, but only DNA evidence (such as from scats) would offer an opportunity to differentiate this animal from other leopards. Although remaining skeptical, we hope that, given the potential importance for conservation, further investigation will deepen the matter."
(Rossi et al., 2020:409)
Following the reported rediscovery of the Zanzibar leopard, student Andrew Weier visited Unguja to investigate the claim. Rather than confirm it, he found no evidence for the real Zanzibar leopard, and instead found that other animals had started to be referred to as leopards (Weier, 2019). To me, this indicates that the real Zanzibar leopard is subconsciously acknowledged as extinct by the local people, which leaves a "leopard" hole which the locals are now filling with other species. A similar occurrence happened after the extinction of the Dodo (Raphus cucullatus) in the mid-17th century, when its name was transferred to the unrelated Red rail (Aphanapteryx bonasia) (Cheke, 2006). The Abstract from his project reads:
“In this project, the modern narrative of the Zanzibar leopard was studied in Jozani-Chwaka Bay National Park. Game cameras were installed and various locations around the national park to try and gather primary evidence that supported the existence of a leopard population in the forest. In addition, local community members were interviewed about recent leopard related activities. No biological evidence was gathered that supported the existence of the Zanzibar leopard and information collected from interviews indicated a potential shift in which other organisms are considered leopards. Recommendations were made for future research to be able to better understand the complex meaningfulness of the narrative of the Zanzibar leopard and how the narrative shifts over time."
(Weier, 2019)
There has been no update on the status of the Zanzibar leopard provided by Galante for five years as of late 2023, and nobody else has been able to capture evidence of it in the last few decades. The island is very small, and it is almost inconceivable that a leopard population could evade documentation for decades despite many people trying.
Special #1: Pondicherry shark (Carcharhinus hemiodon)
The first criterion for a target species should be that it is actually missing, and hasn’t already been rediscovered. Unfortunately, the narrative spun as part of Galante’s first Extinct or Alive special, aired during Shark Week 2019, is that the species hadn’t been recorded since the 1970’s. And therefore, while Galante himself admits that he didn’t personally rediscover the species, he nonetheless attributes that honour to his wife. In his team’s research he has seemingly based the date of the last record on the IUCN RedList entry for the species (Compagno et al., 2003). Which is a strange move since he disregards that same source when it says that species he targets as missing have actually been collected many times in recent years and hence are not missing in the first place (see Special #2).
Online news stories that would have been available via Google to Galante report that the Pondicherry shark was recorded several times during the mid-to-late 1980’s, and was photographed in 2016 (Rodrigo, 2016). While a specimen was caught in 2018, with the article on the catch stating that it was also recorded in 2007 as well (Sankar, 2018). So either Galante has simply not done his research, or he has chosen to ignore local (and) scientific knowledge. This poor target choice is all the more acute since there is a systematic failure to target smaller species, particularly invertebrates. Galante would rather, it seems, search for a large animal that is not even missing, than shine a light on one of the many missing invertebrates.
In 2020, the IUCN RedList re-assessed the species, which was published the following year (Kyne et al., 2021). Rather than accepting the 2019 rediscovery of the species as promoted by Galante and the show, the IUCN assessment went in the opposite direction by rejecting the 1979 record of the previous assessment from 2003 (Compagno et al., 2003), and instead notes that all known museum specimens were collected prior to 1960. With claimed reports since then (i.e. 1979, 1990's, 2000), including some photographs, being misidentifications or otherwise lacking support:
"The contemporary range of this species is poorly defined and museum specimens were collected pre-1960. There are reports from 1979, the 1990s, and 2000, but none of these could be verified. Its identification is problematic, and it is easily confused with a number of other Carcharhinus species (for example, recent putative records from Sri Lanka)."
...
"Misidentification and misreporting through confusion with other carcharhinids are ongoing issues, although there now exists reasonable capacity and knowledge across its historic range to correctly identify Pondicherry Shark if it persists. The increasing use of genetic barcoding, including for dried fins or other body parts, has also increased the likelihood of detection of rare species. Despite this, no samples that would be attributable to this species have been detected using such barcoding techniques."
(both quotes from Kyne et al., 2021)
Season 2, Episode 1: Fernandina Island tortoise (Chelonoidis phantasticus)
The Fernandina Island tortoise (Chelonoidis phantasticus) was described in 1907 based upon a specimen collected the previous year (Van Denburgh, 1907). However, it had never been seen since, partly because nobody had specifically looked for it. And even though there were occasional signs over the years that tortoises still inhabited the island, it was assessed as possibly extinct by the IUCN (Rhodin et al., 2017). Subsequently, an expedition was planned to search for the species on Fernandina Island to confirm its persistence. A television crew tagged along, but when an individual tortoise was found by ranger Jeffreys Málaga, the host of the television show, who had nothing to do with the planning, seized the opportunity and took public the credit for the find. The specific details of this highly unethical behaviour by Forrest Galante are given by (Wight, 2020). Although I have not read his new book, 'Still Alive: A Wild Life of Rediscovery' (2021), it appears that he now does credit ranger Jeffreys Málaga with having found the tortoise. But the audience for a popular television show is far more than for a non-fiction book, so that the damage done was not nearly undone and almost certainly will never be.
Season 2, Episode 2: Miller's grizzled langur (Presbytis canicrus) (added 28 January 2023)
Miller's grizzled langur (Presbytis canicrus) had already been rediscovered in 2011 (Lhota et al., 2012), long before the episode in which Galante is portrayed as having rediscovered it was made. Photos of the species shown on the latest Joe Rogan podact episode to feature Galante (#1927) were taken by the original rediscovery team, as pointed out by one of their members, Dr. Brent Loken, a scientist with more than 8,500 citations:
https://twitter.com/brentloken/status/1617502125143085056
Season 2, Episode 3: Cape lion (Panthera leo melanochaitus)
The Cape lion (Panthera leo melanochaitus) has traditionally been considered one of the earliest megafaunal victims of the European colonisation of southern Africa (last record 1858). Along with the Bluebuck (Hippotragus leucophaeus) (last record c. 1800) and Cape warthog (Phacochoerus aethiopicus aethiopicus) (last record c.1865). However, a revised taxonomy of the entire family Felidae concluded that the Cape lion is in fact extant in virtue of an extant subspecies being a synonym of it (Kitchener et al., 2017). The same conclusion was reached about the Barbary lion (Panthera leo leo). Thus even if you don’t accept the proposed taxonomic changes, it is more likely that the status of the Cape lion will be resolved in the laboratory rather than the field.
Therefore, again it is a wasted opportunity to search for an invertebrate, such as the equal largest moth known from New Zealand, with an almost 15cm (6 inch) wingspan. Buller’s moth (Aoraia mairi) was collected only once, in the summer of 1867, by Sir Walter Buller and his brother-in-law, Captain Gilbert Mair, in the Ruahine Range of the North Island (Meads, 1990). Sadly the specimen is now believed lost.
But the most important point related to this episode is one of Galante’s claims as a guest on Joe Rogan’s podcast #1403:
Therein, Galante states that a particularly large lion with a black mane that his team tranquilised and took a genetic sample from had DNA with a 14% discrepancy from other southern African samples. This may not seem significant, but consider the oft-repeated claim that Chimpanzees and humans share somewhere between 96-98% of their DNA. So this would mean that the lion sampled is not merely a subspecies or even species of lion but a new genus entirely separate from Panthera (which includes the lion, tiger, leopard, jaguar and snow leopard). But instead of talking about a new genus of big cats, which would be one of the greatest zoological discoveries of the 21st century, he talks about a “14% discrepancy” which demonstrates that he pretends to understand genetics3.
Season 2, Episode 7: Rio Apaporis caiman (Caiman crocodilus apaporiensis)
I have nothing new to add here to the fantastic investigative journalism piece by Andrew J. Wight (Wight, 2020). Galante tried to steal credit for the rediscovery of the Rio Apaporis caiman (Caiman crocodilus apaporiensis) from Sergio Balaguera-Reina, who had already announced it. Moreover, Galante has not been forthcoming with the support he promised scientist Sergio Balaguera-Reina in return for helping Galante make the episode a success:
“According to Balaguera-Reina, Galante promised to help fund the Colombian scientist’s caiman research. “But I never saw any of it,” Balaguera-Reina said. Then Galante vanished off Balaguera-Reina’s radar.”
(Wight, 2020)
Special #2: Whitetip weasel shark (Paragaleus leucolomatus) and Flapnose houndshark (Scylliogaleus quecketti)
The latest episode of Galante’s show, technically a Shark Week special in the spirit of the previous year’s, features the rediscovery of not one, not two, but three species! An amazing feat since two, and probably all three, of those species were never missing in the first place. In the first special Galante did for the show, he stated that the Pondicherry shark (Carcharhinus hemiodon) hadn’t been seen since the 1970’s. The only reasonable conclusion to draw as to his source for this claim is the IUCN’s RedList which is available online.
Yet for this latest episode he not merely dispensed with this legitimate though fallible source, he deliberately ignored it. The two species he implies he helped rediscover are the Whitetip weasel shark (Paragaleus leucolomatus) and Flapnose houndshark (Scylliogaleus quecketti), which he claims were last recorded 36 years ago (1984) and almost 120 years ago, respectively (Georgiou, 2020). Yet the respective last records for those two species are later than 2007 (Pollom et al., 2020) and later than 2011 (Pollom, 2019). Galante is now simply making up the last records of (sub)species.
Conclusion
Forrest Galante has a large social media following, and one observation that his fans keep making is that he is truly passionate about wildlife. But passion is disconnected from any reliable metric of someone's competence. You cannot infer how much someone actually knows about a subject by their passion for it. And the evidence is clear that he makes basic errors, either cannot do proper research to ascertain the correct conservation status/taxonomic validity of taxa or doesn't care to, and partakes in unethical behaviour by stealing and attempting to steal credit from legitimate researchers. But the greatest evidence that he is not a competent biologist is his singular failure to have followed through with any of these alleged rediscoveries by publishing them in a peer-reviewed scientific journal. He thus maximises his public image by constructing a rediscovery narrative that the public are not likely to question, while easing the burden upon himself by not putting it out there for the scientific community to properly vet.
The most insidious thing about his impact is the projected general sense that he has rediscovered so many animals that scientists have systematically failed, over a much longer period, to find. Forrest and his TV crew only search briefly and yet have a far greater rate of success. And therefore conservationists proper are at best ignorant, and at worst incompetent. And that has profound ramifications for how members of the public decide to spend or donate their money. The generosity of the public in donating to conservation and environmental causes is not something to be taken for granted at the best of times. People work hard for their money, and have the right to spend it as they see fit. While the greed of humanity as a whole has meant that there are so many species, subspecies and populations in danger of extinction that the resources required to adequately to save them all will almost certainly not be forthcoming. So whatever funds can be secured are all the more important.
Television shows are for entertainment purposes and therefore do not always have technical accuracy in mind when presenting their material. And of course such shows are the products of many people not just the host. But whatever the constraints that were placed upon him when agreeing to do the show, no such conditions would be worth compromising for to a genuine conservationist if they resulted in such a distorted view of conservation biology as actually resulted from the show. The concept of looking for a different putatively extinct (sub)species each week is a novel and laudable idea, but it would have been infinitely served better if there had been no falsely claimed rediscoveries. It would ultimately have been realistic, because searching for missing organisms is hard, very hard, and often takes lots and lots of planning and searching around for funding.
Within science, the bread and butter of communication and influence are peer-reviewed papers that are referreed by two reviewers that are anonymous to the paper's authors to help ensure a high standard of output. This process can take a long time, and currently my Twitter feed is increasingly consisting of journal editors and authors lamenting the difficulty in finding reliable peer-reviewers, meaning that papers can take years to have published. But while this process is unfortunately drawn out, it is not without end. And yet Forrest Galante still has not published any of his alleged rediscoveries in a vetted peer-reviewed scientific paper. Indeed, when he is asked about updates regarding any of these species he invariably states that he has passed off the species to another researcher/group or has not heard anything new. This does not strike one as the approach of a legitimate individual who is intersted in conserving species, but rather that of an imposter who is only interested in maximising his own public image.
If I rediscovered even a single species, I would want every update on its status imaginable. To simply walk away from it shows a lack of real interest in its fate. Forrest Galante is then, ultimately, a charlatan.
Postscript:
Two weeks after initially writing this article I found the following quote from the "talk" section [archived version] of the Wikipedia page on Forrest Galante:
"Questionable removals
Two sentences that cited criticism of Galante were removed three times from IPs near Galante's residence & Animal Planet offices. The source, Undark, does not attempt to appear neutral on the topic of its criticism, but does have editorial control on pieces published and cites significant figures in relevant fields on the topic. The sentence referencing the Galapagos episode in particular cites an extremely relevant source that was present at the filming. Especially considering much of the article reading like an advertisement, this does not seem like good-faith editing. The removals appear to be based on hiding criticism of Galante rather than removing unreliable or hidden-bias sources."
Notes
1 He has six spearfishing world records according to his Facebook page.
2 Mike, along with Gareth Linnard and myself, co-(re)discovered three historical films of the species, including the latest known footage of the species (March-April 1935), which went viral around the world in 2020. One of which Forrest himself shared on Instagram.
3 If the 14% "discrepancy" relates only to the mitogenome then the genetic distance would be considerably smaller, but still extremely significant, and would still sustain the overall criticism.
References
Buckley-Beason V. A., Johnson W. E., Nash W. G., Stanyon R., Menninger J. C., Driscoll C. A, Howard J. G., Bush M., Page J. E., Roelke M. E., Stone G., Martelli P. P., Wen C., Ling L., Duraisingam R. K., Lam P. V. and O’Brien S. J. (2006). Molecular evidence for species-level distinctions in clouded leopards. Current Biology 16(23): 2371-2376. https://doi.org/10.1016/j.cub.2006.08.066
Cameron, Robert A. D., Teixeira, Dinarte, Pokryszko, Beata, Silva, Isamberto and Groh, Klaus. (2021). An annotated checklist of the extant and Quaternary land molluscs of the Desertas Islands, Madeiran Archipelago. Journal of Conchology 44(1): 53-70.
Cheke, Anthony S. (2006). Establishing extinction dates - the curious case of the Dodo Raphus cucullatus and the Red Hen Aphanapteryx bonasis. Ibis 148: 155-158. https://doi.org/10.1111/j.1474-919X.2006.00478.x
Compagno, L. J. V., White, W. and Fowler, S. (SSG Australia & Oceania Regional Workshop, March 2003). (2003). Carcharhinus hemiodon. The IUCN Red List of Threatened Species 2003: e.T39369A10185838. https://dx.doi.org/10.2305/IUCN.UK.2003.RLTS.T39369A10185838.en. Downloaded on 16 August 2020.
Fredriksson, Gabriella M. (2005). Predation on sun bears by reticulated python in East Kalimantan, Indonesian Borneo. The Raffles Bulletin of Zoology 53(1): 165-168.
Georgiou, Aristos. (2020). Shark Not Seen for Over 100 Years Rediscovered in 'Land of the Lost'. Newsweek, 10 August, available at: https://www.newsweek.com/shark-not-seen-over-100-years-rediscovered-1524094 [Accessed 16 August 2020]
Gerlach, Justin et al. (2006). The first substantiated case of trans-oceanic tortoise dispersal. Journal of Natural History 40(41-43): 2403-2408.
Goldman, Helle V. and Walsh, Martin. (2018). “The Zanzibar Leopard” on Animal Planet. Blogpost, available at: http://zanzibarleopard.blogspot.com/2018/06/the-zanzibar-leopard-on-animal-planet.html [Accessed 15 August 2020]
Hendrich, Lars, Manuel, Michael and Balke, Michael. (2019). The return of the Duke—locality data for Megadytes ducalis Sharp, 1882, the world's largest diving beetle, with notes on related species (Coleoptera: Dytiscidae). Zootaxa 4586(3): 517-535. [Abstract]
Jones, Menna. (2023). Introduction: The thylacine in Australian ecosystems, pp. xxv-xxviii. In: Holmes, Branden and Linnard, Gareth (eds.). Thylacine: The History, Ecology and Loss of the Tasmanian Tiger. Clayton South, Victoria: CSIRO Publishing. 240 pp.
Kitchener, A. C., Beaumont, M. A. and Richardson, D. (2006). Geographical variation in the clouded leopard, Neofelis nebulosa, reveals two species instead of one. Current Biology 16(23): 2377-2383. https://doi.org/10.1016/j.cub.2006.10.066
Kitchener, A. C., Breitenmoser-Wursten, Ch., Eizirik, E., Gentry, A., Werdelin, L., Wilting, A., Yamaguchi, N., Abramov, A. V., Christiansen, P., Driscoll, C., Duckworth, J. W., Johnson, W., Luo, S.-J., Meijaard, E., O, Meijaar, P., Sanderson, J., Seymour, K., Bruford, M., Groves, C., Hoffmann, M., Nowell, K., Timmons, Z. and Tobe, S. (2017). A revised taxonomy of the Felidae. The final report of the Cat Classification Task Force of the IUCN/SSC Cat Specialist Group. Cat News Special Issue 11: 1-80.
Kyne, P.M., Jabado, R.W., Akhilesh, K.V., Bineesh, K.K., Booth, H., Dulvy, N.K., Ebert, D.A., Fernando, D., Khan, M., Tanna, A. & Finucci, B. (2021). Carcharhinus hemiodon. The IUCN Red List of Threatened Species 2021: e.T39369A115736695. https://dx.doi.org/10.2305/IUCN.UK.2021-1.RLTS.T39369A115736695.en. Accessed on 30 July 2022.
Lhota, S., Loken, B., Spehar, S., Fell, E., Pospěch, A. and Kasyanto, N. (2012). Discovery of Miller's Grizzled Langur (Presbytis hosei canicrus) in Wehea Forest Confirms the Continued Existence and Extends Known Geographical Range of an Endangered Primate. American Journal of Primatology 74(3): 193-198. https://doi.org/10.1002/ajp.21983
Meads, Michael J. (1990). Forgotten fauna: the rare, endangered and protected invertebrates of New Zealand. DSIR Publishing: Wellington.
Miththapala, Sriyanie, Seidensticker, John and O'Brien, Stephen J. (1996). Phylogeographic Subspecies Recognition in Leopards (Panthera pardus): Molecular Genetic Variation. Conservation Biology 10(4): 1115-1132.
Nowak, R. M. (1995). Another look at wolf taxonomy, pp. 375-397. In: Carbyn, L. N., Fritts, S. H. and Seip, D. R. (eds.). Proceedings of the second North American symposium on wolves. Edmonton, Alberta: Canadian Circumpolar Institute, University of Alberta.
Patterson, Robert. (1973). Why Tortoises Float. Journal of Herpetology 7(4): 373-375. [First page]
Patton, A. H., Lawrance, M. F., Margres, M. J., Kozakiewicz, C. P., Hamede, R., Ruiz-Aravena, M., ... and Storfer, A. (2020). A transmissible cancer shifts from emergence to endemism in Tasmanian devils. Science 370(6522): eabb9772. https://doi.org/10.1126/science.abb9772
Pocock, Reginald I. (1932). The leopards of Africa. Proceedings of the Zoological Society of London II 1932: 543-591.
Pollom, R., Bennett, R., Ebert, D.A., Gledhill, K., McCord, M.E. & Kyne, P.M. (2020). Paragaleus leucolomatus. The IUCN Red List of Threatened Species 2020: e.T161639A124519483. https://dx.doi.org/10.2305/IUCN.UK.2020-2.RLTS.T161639A124519483.en. Downloaded on 15 August 2020.
Pollom, R., Da Silva, C., Ebert, D. A. and Fennessy, S. (2019). Scylliogaleus quecketti. The IUCN Red List of Threatened Species 2019: e.T39360A124406361. Downloaded on 16 August 2020.
Provost, Stephen H. (20 May, 2016). Animal Planet’s ‘Extinct or Alive’ TV special stars Coast Union grad. The Tribune (online), available from: https://www.sanluisobispo.com/news/local/community/cambrian/cambrian-arts-events/article78878307.html [Accessed 15 August 2020]
Rabinowitz, Alan. (2005). Jaguars and livestock: living with the world’s third largest cat, pp. 278-285. In: Woodroffe, R., Thirgood, S. and Rabinowitz, A. (eds.). People and Wildlife, Conflict or Coexistence? Cambridge, UK: Cambridge University Press.
Rhodin, A. G. J., Gibbs, J. P., Cayot, L. J., Kiester, A. R. and Tapia, W. (2017). Chelonoidis phantasticus (errata version published in 2018). The IUCN Red List of Threatened Species 2017: e.T170517A128969920. https://dx.doi.org/10.2305/IUCN.UK.2017-3.RLTS.T170517A1315907.en. Downloaded on 16 August 2020.
Rivas, Jesús Antonio. (2000). The life history of the green anaconda (Eunectes murinus), with emphasis on its reproductive biology. PhD dissertation, The University of Tennessee.
Rossi, L., Scuzzarella, C. M. and Angelici, F. M. (2020). Extinct or Perhaps Surviving Relict Populations of Big Cats: Their Controversial Stories and Implications for Conservation, pp. 393-417. In: Angelici F., Rossi L. (eds.). Problematic Wildlife II. Cham: Springer.
Sankar, K. N. Murali. (2018). ‘Pondicherry shark’ spotted near Kakinada. The Hindu, 10 September, available at: https://www.thehindu.com/news/national/andhra-pradesh/pondicherry-shark-spotted-near-kakinada/article24910627.ece [Accessed 16 August 2020]
Setiawan, A., Nugroho, T. S., Djuwantoko (mononym) and Pudyatmoko, S. (2009). A survey of Miller’s grizzled surili, Presbytis hosei canicrus, in East Kalimantan, Indonesia. Primate Conservation 24: 139–143. https://doi.org/10.
Smith, J. L. B. (1956). Old Fourlegs: The Story of the Coelacanth. Longmans Green.
Sun, Xin et al. (2023). A genomic exploration of the possible de-extirpation of the Zanzibar leopard. bioRxiv preprint. https://doi.org/10.1101/2023.07.25.550323
Swingler, Helen. (2023, July 21). PhD graduand clocks vast distances on foot collecting 150 taxa, including 50 undescribed species. University of Cape Town (news story). Available at: https://www.news.uct.ac.za/news/graduation/2023/july/-article/2023-07-21-phd-graduand-clocks-vast-distances-on-foot-collecting-150-taxa-including-50-undescribed-species [Accessed 12 October 2023]
Uphyrkina, O., Johnson, E. W., Quigley, H., Miquelle, D., Marker, L., Bush, M. and O'Brien, S. J. (2001). Phylogenetics, genome diversity and origin of modern leopard, Panthera pardus. Molecular Ecology 10(11): 2617-2633.
Van Denburgh, J. (1907). Expedition of the California Academy of Sciences to the Galapagos Islands, 1905-1906. I. Preliminary descriptions of four new races of gigantic land tortoises from the Galapagos Islands. Proceedings of the California Academy of Sciences 1(4): 1-6.
Walsh, Martin T. and Goldman, Helle V. (2012). Chasing imaginary leopards: science, witchcraft and the politics of conservation in Zanzibar. Journal of Eastern African Studies 6(4): 727-746. [Abstract]
Weier, Andrew. (2019). Leopards Are Good to Think With: Spotting the Zanzibar Leopard in Jozani Forest. Unpublished paper.
Weinberg, Samantha. (2000). A Fish Caught in Time: The Search for the Coelacanth. HarperCollins Publishers. 256 pp.
Wight, Andrew J. (2020). In the Bombast of an American TV Host, Colonial Science Lives On. Undark, 3 April, available at: https://undark.org/2020/03/04/colombia-reptile-parachute-science-forrest-galante/ [Accessed 16 August 2020]
Wild, Sarah. (2019, November 5). Cape student rediscovers third “extinct” plant species – and expects to find more. Business Insider South Africa, available at: https://www.news24.com/news24/bi-archive/extinct-plant-species-sweet-pea-stellenbosch-student-nature-2019-11 [Accessed 15 August 2020]
Woods, Gregory. (2023). Prologue, pp. xxii-xxiv. In: Holmes, Branden and Linnard, Gareth (eds.). Thylacine: The History, Ecology and Loss of the Tasmanian Tiger. Clayton South, Victoria: CSIRO Publishing. 240 pp.
How Smart Were Neanderthals? Or, How Dumb Are We?
By Branden Holmes
Introduction
The long prevailing view of Neanderthals Homo neanderthalensis (King, 1864) as brutish, slow-witted, technologically inferior, cavemen (in the pejorative sense) is hopefully now behind us. However, it should never have taken 3-ply fibres recovered from a Neanderthal site (Hardy et al., 2020), to take the most recent example published, to infer that Neanderthals were about as smart as we are. Such technological finds, due to their fragile nature, are very rarely preserved, and offer a rather direct look at Neanderthal culture. And in that sense are truly marvellous, reminding us that we are, or at least were not, intellectually unique in the universe. Not to mention the potential for such intelligence to evolve again in the future, on Earth or elsewhere in the universe.
But leveraging such discoveries should never have been needed. In fact one does not even need to appeal to the reality that for thousands upon thousands of years Neanderthals lived alongside the other top predators of Eurasia, and wherever else they may have lived but have not yet been found: Cave Bears (Ursus spelaeus), Cave Lions (Panthera spelaea), Scimitar-toothed cats (Homotherium latidens), etc. etc. Any dim-witted biped stumbling around Europe during the Late Pleistocene (c.126ka-11.7ka) would simply not have survived, in an environment which makes today's Africa look rather tame. Of course, the comparison with today's Africa is somewhat unfair, since Africa itself has lost a great deal of its megafauna too (Faith, 2014). A fact that has only relatively recently risen to prominence. Including the spectacular discovery of a gigantic partial skull of a lion, equal in size to the largest Cave lions of Eurasia, and much larger than any hitherto found in Africa (Manthi et al., 2018). Suggestive of a previously unknown taxon, and not merely palaeoclinal variation (Ibid.). And which may very well have survived into the Late Pleistocene (c.126ka-11.7ka), since stratigraphic extent cannot be inferred based upon a single record.
Moreover, it is simply not true that only large carnivores pose a mortal threat. It is not merely a pun to suggest that hunting a Mammuthus primigenius would be a mammoth task. Not to mention the many other, much smaller, deadly creatures that inhabit the seven continents. Nor should it take the realisation that we Homo sapiens weren't exactly brimming with brilliant ideas. Evidence suggests we only began seriously farming around 11,500 years ago, long after the last evidence for Neanderthals at 28 ± 3 ka on the Rock of Gibraltar (Muñiz et al., 2019). And today's humans are hardly a paradigm of intellectual superiority, just look on YouTube. Or to take a truly contemporary example, people who get tested for COVID-19 but do not self isolate while awaiting the results. Especially when their coughing turns into a coffin for somebody else. No, the basic reason to suppose that Neanderthals were basically as smart as we are is the burden of differentiation which taxonomy, and to a lesser extent nomenclature, is predicated upon.
Taxonomy and Nomenclature
Given the current coronavirus pandemic, the medical term 'diagnosis' is all too familiar. And just as in non-medical contexts, the term indicates a problem or malady. Whereas in the unique sense that it is deployed in taxonomy and nomenclature, it means virtually the opposite. To "diagnose a species" is essentially to state what is healthy and robust about it. That is, how it differs from previously known species. This burden of proof of demonstrating relevant (consistent) difference is derived from the fact that neither commonality nor some types of difference will do the job. For example, knowing that two organisms are both green is compatible with them belonging to the same species, and with them belonging to different species. Likewise, two organisms being different colours is compatible with them belonging to different species, and to them belonging to the same species. The difference in colour may simply be ontogentically related, or due to sexual dimorphism.
Of course, species are categorised in terms of higher taxonomy too. For example, at the family (or familial) level or at the genus level. As far as we're aware, life on Earth has a single origin, possibly 4 billion years ago. So since Homo neanderthalensis and Homo sapiens are categorised in the same genus, which evidence suggests arose only around 2-2.5 million years ago (Dunsworth, 2010), the two species share virtually all of their evolutionary history together. And so it is entirely natural for them to be extremely similar in many respects, including with respect to intelligence level. The differences between them are then relatively minor compared with the similarities. And so unless one can diagnose the two species based upon differential intelligence, then it is perfectly reasonable to assume that their intelligence is comparable, or at least to remain agnostic about it until further discoveries are made. Either way, it makes no logical sense to suppose that we humans are somehow arbitrarily unique, even amongst our closest living and extinct relatives.
Conclusion
Neither the fossil remains, nor the cultural items, which have been found to belong to the Neanderthals give any indication of significantly lesser intelligence than us. On the face of it this is no surprise, since we share so much evolutionary history, and have only relatively recently diverged. And yet there is a pervasive belief, and not merely among scientists, that our species is somehow intellectually superior to the rest of the animal kingdom. Nothing could be further from the truth. We are of comparable intelligence to Neanderthals, and hence asking "how smart were Neanderthals?" is the same as asking "how dumb are we?". And vice versa.
References
Dunsworth, Holly M. (2010). Origin of the Genus Homo. Evolution: Education and Outreach 3: 353-366.
Faith, J. Tyler. (2014). Late Pleistocene and Holocene mammal extinctions on continental Africa. Earth-Science Reviews 128: 105-121.
Hardy, B. L., Moncel, M.-H. et al. (2020). Direct evidence of Neanderthal fibre technology and its cognitive and behavioral implications. Scientific Reports 10: 4889.
Manthi, Fredrick K., Brown, Francis H., Plavcan, Michael J. and Werdelin, Lars. (2018). Gigantic lion, Panthera leo, from the Pleistocene of Natodomeri, eastern Africa. Journal of Paleontology 92(2): 305-312.
Muñiz, Fernando et al. (2019). Following the last Neanderthals: Mammal tracks in Late Pleistocene coastal dunes of Gibraltar (S Iberian Peninsula). Quaternary Science Reviews.
Local Rediscoveries
If an extirpated species or subspecies is rediscovered within that area, provided recolonisation/translocation from another area can be ruled out, then a genuine local rediscovery has real import for conservation biology. If a large carnivore is thought to have been extirpated from a small island, but is later rediscovered there, then its having genuinely gone undetected for a long period (despite methodologically appropriate survey methods) demonstrates that the classification of "extirpated" was premature. It is entirely irrelevant that individuals of that taxon survive elsewhere (since we have theoretically ruled out recolonisation/translocation), and therefore a population went genuinely undetected despite surveys for a significant period.
Afghanistan
Algeria
Ben Si Said, Z. and Rebbas, K. (2022). Rediscovery of Ophrys omegaifera subsp. hayekii (H. Fleisch. & Soó) and Dactylorhiza markusii (Tineo) H. Baumann & Künkele Kreutz in the region of Tizi Ouzou (Great Kabylia, Algeria). Journal of EcoAgriTourism 18(2): 47-53.
Chaker, Amina et al. (2023). New data on lichen flora of the Edough Peninsula in north-eastern Algeria. Botanica 29(1): 9-20. [Fuscopannaria leucosticta]
Chelli, Abdelmadjid, Moulaï, Riadh and Djemai, Abdelhadi. (2020). Does the Tichi Haf dam construction affect dragonfly and damselfly (Odonata : Insecta) assemblages of the Boussellam watercourse (central north Algeria)? A preliminary study. Zoology and Ecology 30(1): 37-47.
Glöer, P. and Ramdini, R. (2019). Hippeutis complanatus Linnaeus, 1758 in Algeria, refound after more than 150 years. Ecologica Montenegrina 22: 226-227. https://doi.org/10.37828/em.2019.22.18.
Khelifa, Rassim and Zebsa, Rabah. (2018). Rediscovery of the regionally critically endangered dragonfly Lindenia tetraphylla in Northeast Algeria after 170 years of apparent absence (Odonata: Gomphidae). Notulae odonatologicae 9(2) 2018: 50-54.
Ramdini, R., van Damme, D., Sadouk, G. and Medjdoub-Bensaad, F. (2020). Rediscovery of Armiger crista (Linnaeus, 1758) (Gastropoda, Planorbidae) in Algeria. Biodiversity Journal 11(4): 821-824. https://doi.org/10.31396/Biodiv.Jour.2020.11.4.821.824
Slimani, S. et al. (2020). New reports of the Argentine ant Linepithema humile (Mayr, 1868) (Hymenoptera: Formicidae) in Algeria. Ukrainian Journal of Ecology 10(5): 248-252. doi:10.15421/2020_239
Argentina
Etchepare, E. G.; V. H. Zaracho. (2009). Serpentes, Colubridae, Taeniophallus poecilopogon (Cope, 1863): Rediscovery in Corrientes, Argentina, and natural history. Check List 5(4): 770-772.
Leuchtenberger, Caroline et al. (2021). Hope for an apex predator: giant otters rediscovered in Argentina. Oryx 55(6): 810-811.
Nenda, S. J.; D. O. Di Pietro. (2009). Serpentes, Dipsadidae, Echinantherini, Taeniophallus poecilopogon (Cope, 1863): Rediscovery in Argentina. Check List 5(3): 503-506.
Pardiñas, Ulyses F. J. et al. (2008). Rediscovery of Juliomys pictipes (Rodentia: Cricetidae) in Argentina: emended diagnosis, geographic distribution, and insights on genetic structure. Zootaxa 1758(1): 29-44. [Abstract]
Schejter, Laura, Cristobo, Javier and Ríos, Pilar. (2022). Rediscovering Fibulia myxillioides (Burton, 1932) (Porifera, Poecilosclerida) in the SW Atlantic Ocean. Panamjas 17: 16. doi.org/10.54451/PanamJAS.17.1.16
Armenia
Arakelyan, Marine, Ananian, Vasil and Petrosyan, Ruzanna. (2019). Rediscovery of Chernov’s skink (Ablepharus chernovi Darevsky, 1953) in Armenia. Herpetology Notes 12: 475-477.
Rudov, A., Nersesyan, A. and Asatryan, A. (2024). New data on the genus Ferula (Apiaceae) for the flora of Armenia; a new record and a rediscovery. Iran. J. Bot. 30(2): 132-138. [Ferula glabrifolia]
Australia
Aitken, P. F. (1971). Rediscovery of the Large Desert Sminthopsis (Sminthopsis psammophilus Spencer) on Eyre Peninsula, South Australia. Victorian Naturalist 88: 103-111.
Aitken, P. F. (1977). Rediscovery of swamp antechinus in South Australia after 37 years. South Australian Naturalist 52: 28–30. [Antechinus minimus maritimus]
Baynes, Alexander, Chapman, Andrew and Lynam, Antony J. (1987). The rediscovery, after 56 years, of the heath rat Pseudomys shortridgei (Thomas, 1907) (Rodentia: Muridae) in Western Australia. Records of the Western Australian Museum 13(2): 319-322.
Bourke, Patricia M and Willan, Richard C. (2009). 'Anadara granosa' (Mollusca: Bivalvia: Arcidae) Discovered Live in Darwin Harbour, with Implications for Understanding Climate Change in Northern Australia. The Beagle: Records of the Museums and Art Galleries of the Northern Territory 25: 115-118. [Abstract]
Cheal, David. (2010). Velvet Thread-petal 'Stenopetalum velutinum' Rediscovery in Victoria. The Victorian Naturalist 127(1): 19-22. [Abstract]
Doody, J. Sean, Rhind, David, Castellano, Christina M. and Bass, Michael. (2012). Rediscovery of the scaly-tailed possum (Wyulda squamicaudata) in the eastern Kimberley. Australian Mammalogy 34(2): 260-262. [Abstract]
Eldridge, M. D. B. and Close, R. L. (1995). Brush-tailed Rock-wallaby Petrogale penicillata, pp. 383-385. In: Strahan, Ronald (ed.). The Mammals of Australia. Chatswood: Reed Books Australia. [rediscovery in Victoria]
Falconer, Audrey and Burn, Robert. (2010). Rediscovery of the Seaweed Limpet 'Naccula parva' in Victorian Waters. The Victorian Naturalist 127(6): 246-247. [abstract]
Gibb, W. F. (1977). The rediscovery of Hypolimnas antilope (Cramer) (Lepidoptera: Nymphalidae) in Australia. Australian Entomological Magazine 4: 39.
Hammer, Michael P. et al. (2015). Regional extinction, rediscovery and rescue of a freshwater fish from a highly modified environment: The need for rapid response. Biological Conservation 192: 91-100. [Abstract]
Houston, Wayne, Porter, Gary, Elder, Rod, Black, Robert and Sheaves, Marcus. (2004). Rediscovery of yellow chats (Capricorn subspecies) on the Fitzroy River delta central Queensland. The Sunbird: Journal of the Queensland Ornithological Society 34(1): 36-42. [abstract]
Humphreys. W. F.. Brooks, R. D. and Bine. B. (1990). Rediscovery of the palm Livistona alfredii on North West cape Peninsula. Records of the Western Australian Museum 14(4): 647-650.
Jaensch, Roger, Houston, Wayne, Black, Robert, Campbell, Lorelle, McCabe, John, Elder, Rod and Porter, Gary. (2004). Rediscovery of the Capricorn subspecies of yellow chat 'Epthianura crocea macgregori' at Torilla Plain, on the mainland coast of central Queensland. The Sunbird: Journal of the Queensland Ornithological Society 34(1): 24-35. [abstract]
Kerrigan, Raelee, Dixon, Dale and Cowie, Ian. (2003). Ptychosperma macarthurii or P. bleeseri? The Taxonomic Status of P. bleeseri Reconsidered. The Beagle: Records of the Museums and Art Galleries of the Northern Territory 19: 81-86. [abstract]
New, T. R. (2007). The Hemiphlebia Damselfly 'Hemiphlebia mirabilis' Selys (Odonata, Zygoptera) as a Flagship Species for Aquatic Insect Conservation in South-Eastern Australia. The Victorian Naturalist 124(4): 269-272. [abstract]
Warneke, R. M. (1960). Rediscovery of the broad-toothed rat in Gippsland. Victorian Naturalist 77: 195-196. [Mastacomys fuscus]
ACT
Osborne, W. S., K. Kukolic, M. S. Davis and R. Blackburn. (1993). Recent records of the earless dragon Tympanocryptis lineata pinguicolla in the Canberra region and a description of its habitat. Herpetofauna 23(1): 16-25.
Christmas Island
Macquarie Island
New South Wales
Curran, T. J. and Curran, S. R. (2005). Rediscovery of Ooline, Cadellia pentastylis, near Gunnedah: notes on the habitat and ecology of this dry rainforest tree. Cunninghamia 9: 311-316.
Leggett, Keith E. A., Welaratne, Thanuri, Letnic, Michael, McLeod, Steven and Dawson, Terrence. (2017). Rediscovery of the plains mouse (Pseudomys australis) (Rodentia: Muridae) in New South Wales. Australian Mammalogy 40(1): 127-130.
Theischinger, G., Miller, J., Miller, R. & Krogh. M. (2009). Rediscovery of Austrocordulia leonardi (Sydney Hawk) in the suburbia of Sydney. Agrion 13(2): 50-53.
Dasycercus cristicauda (Crest-tailed mulgara)
Boronia deanei (Deane's boronia)
Northern Territory
Braby, Michael F. and Westaway, John O. (2016). Rediscovery of the Spinifex Sand-skipper (Proeidosa polysema) in the Darwin area, Northern Territory. Northern Territory Naturalist 27: 121-125. [Abstract]
Queensland
Barker, Dayana et al. (2022). Rediscovery of Ixodes confusus in Australia with the first description of the male from Australia, a redescription of the female and the mitochondrial (mt) genomes of five species of Ixodes. International Journal for Parasitology: Parasites and Wildlife 18: 1-11. https://doi.org/10.1016/j.ijppaw.2022.03.006
Freeman, A. B., Benham, B., Sebasio, D., Cann, J. and Strevens, W. (2014). Summary of the rediscovery of the Jardine River turtle in Australia: Management implications and recommendations. Brisbane: Department of Environment and Heritage Protection, Queensland Government.
Field, Ashley and Renner, Matthew. (Accepted, 2018). Rediscovered or reconsidered: The presumed Extinct ferns and lycophytes of tropical Queensland, Australia. Australian Systematic Botany. [Abstract] [Oreogrammitis leonardii]
Jerrard, Cyril H. H. (1925). Black-breasted Quail (Turnix melanogaster) in the Burnett district, Queensland. The Emu 25(2): 123. [global rediscovery?]
Huperzia dalhousieana: http://vanishingflora.proboards.com/thread/2091/huperzia-dalhousieana
Monaeses brevicaudatus: https://www.facebook.com/TurnFear2Fascination/photos/?tab=album&album_id=1468149803280911
South Australia
Bates, R. (1981). The rediscovery of Lycopodium deuterodensum Herter in South Australia. S. Austral. Nat. 55(4): 63.
Bates, R. J. and Weber, J. Z. (1990). Orchids of South Australia. South Australia: Government Printer. [Caladenia gladiolata in the Mt. Lofty Ranges]
Carpenter, G. and Horton, P. (1999). Birds, pp. 205-224. In: Robinson, A. C. and Armstrong, D. M. (eds.). (1999). A Biological Survey of Kangaroo Island, South Australia, 1989 & 1990. South Australia: Heritage and Biodiversity Section, Department for Environment, Heritage and Aboriginal Affairs. ["[Psophodes nigrogularis was] rediscovered in South Australia by Hal Crouch on Kangaroo Island in 1967" (p. 221)]
Grund, Roger, Stolarski, Alex and Stevens, Mark. (2018). Rediscovery of Synemon selene (Lepidoptera: Castniidae) in the mid-north of South Australia. Transactions of the Royal Society of South Australia 143(1): 86-91.
Toelken, H. R. (2014). Notes on Hibbertia (Dilleniaceae) 10. Hibbertia hirsuta (subgen. Hemistemma: H. vestita group) does occur in South Australia and Tasmania. Journal of the Adelaide Botanic Gardens 27: 35-39.
Tasmania
Bonham, Kevin J. (1997). Native land snails of King Island and the Hunter Group. The Tasmanian Naturalist 119: 10-22. [Chloritobadistes victoriae]
Bonham, Kevin J. (2008). Rediscovery of Corunastylis nudiscapa (Hook.f.) D.L.Jones & M.A.Clem. in Tasmania. The Tasmanian Naturalist 130: 100-102.
McQuillan, Peter B., Schmidt, Olga, Byrne, Catherine J. and Warren, Margaret. (2024). Rediscovery and systematics of the rare brown-caped carpet moth Chrysolarentia excentrata (Guenée, 1857 [1858]) comb. nov. (Lepidoptera: Geometridae: Larentiinae) in Tasmania. Papers and Proceedings of the Royal Society of Tasmania 158: 79-86.
Skabo, Roy. (2009). Resurrecting Hibbertia rufa (Brown Guineaflower). The Tasmanian Naturalist 131: 12-16.
Wapstra, M., Daniels, G. and White, R. (2020). Two fireweeds back from the dead: the rediscovery of Senecio longipilus and Senecio extensus from the Tasmanian central highlands. The Tasmanian Naturalist 142: 83-89.
Wapstra, M., Duncan, F., Buchanan, A. and Schahinger, R. (2006). Finding a botanical Lazarus: tales of Tasmanian plant species ‘risen from the dead’. The Tasmanian Naturalist 128: 61-85. [Senecio campylocarpus, Bossiaea obcordata, Lycopus australis, Mentha australis, Myosurus australis, Vittadinia australasica oricola]
Wapstra, M., French, B., Tyquin, V. & Skabo, R. (2011). From presumed extinct to probably secure? The resurrection and ongoing management of Hibbertia rufa (brown guineaflower) in north-east Tasmania. Tasforests 19: 54-70.
McLenna, April. (2021, 20 October). New Holland Mouse, thought to be extinct, rediscovered on Tasmania's Flinders Island. ABC News (online), available at: https://www.abc.net.au/news/2021-10-20/native-new-holland-mouse-found-on-flinders/100553568 [Accessed 11 November 2021]
Victoria
Anon. (1995). On the rediscovery of Thelymitra merraniae at Moggs Creek. Native Orchid Soc. S. Austral. J. 19(2): 18-19.
Barker, W. R. (1987). Taxonomic studies in Euphrasia L. (Scrophulariaceae). V. New and Rediscovered taxa, typifications, and other notes on the genus in Australia. J. Adelaide Bot. Gard. 10(2): 201-221.
Beardsell D, Beardsell C. 1986 Rediscovery of Pterostylis woollsii R.D. Fitzgerald in Victoria after half a century. Victorian Nat., 103. (5): 140-141.
BRUNNER, H. & BERTUCH, I.D., 1976: The Broadtoothed rat still in Sherbrooke Forest. A successful search for Mastacomys fuscus Thomas. Vict. Nat. 93: 55-56.
Burns, A.N., 1955. Rediscovery of a “Living Fossil” Damsel-fly in Victoria. Victorian Naturalist 72: 116-117. [Hemiphlebia mirabilis]
Dunn, Kelvyn L. (2020). Rediscovery of the Rayed Blue, Candalides heathi (Cox 1873) (Lepidoptera: Lycaenidae) on Mount Buffalo, Victoria. Calodema, 750: 1-27.
Falconer, Audrey and Burn, Robert. (2010). Rediscovery of the Seaweed Limpet 'Naccula parva' in Victorian Waters. The Victorian Naturalist 127(6): 246-247. [Abstract]
Field, R. P. (1978). Rediscovery of Pseudodipsas brisbanensis in Victoria. Victorian Entomologist 8: 5-7.
Littlejohn, M. and Martin, A. A. (1967). The rediscovery of Heleioporus australiacus (Shaw) [Anura: Leptodactylidae] in eastern Victoria. Proceedings of the Royal Society of Victoria 80(1): 31-36.
Psophodes leucogaster (White-bellied whipbird)
Bearded flatsedge (Cyperus squarrosus)
Western Australia
Baggs, J. W. (1953). Re-discovery of the Ground Parrot at the Bow River. Western Australian Naturalist 3(8): [pagination?].
Harvey, Judith. (2017). Forget-me-not, not forgotten any more: rediscovery of Myosotis australis on Rottnest Island. Landscope [2017]: 36.
Mesembriomys gouldii gouldii (Black-footed tree rat)
O'Donoghue, Mike and Atkins, Ken. (2000). Lost Jewels in the Bush. Landscope 15(3): 23-27. [Lepidium aschersonii]
Petrogale concinna (Nabarlek): https://www.australiangeographic.com.au/news/2018/01/australia%E2%80%99s-second-smallest-rock-wallaby-rediscovered-in-wa
Austria
Hödl, Walter and Eder, Erich. (1996). Rediscovery of Leptestheria dahalacensis and Eoleptestheria ticinensis (Crustacea: Branchiopoda: Spinicaudata): an overview on presence and conservation of clam shrimps in Austria. Hydrobiologia 318: 203-206.
Wanzenböck, J. and Spindler, T. (1995). Rediscovery of Umbra krameri Walbaum, 1792, in Austria and subsequent investigations. Annalen des Naturhistorischen Museums in Wien 97B: 450-457.
Umbra krameri (Wanzenböck & Spindler, 1995)
Bangladesh
Alam, A. B. M. Sarowar et al. (2016). Rediscovery of the Critically Endangered Slender-billed Vulture Gyps tenuirostris in Bangladesh. BirdingASIA 25: 69-70.
Hasan, Mohammad Tarikul and Uddin, Mohammad Zashim. (2022). Rediscovery of Potentilla supina L. (Rosaceae) in Bangladesh. Bangladesh J. Plant Taxon. 29(2): 437-440. https://doi.org/10.3329/bjpt.v29i2.63538
Hossain, Monwar. (2014). Rediscovery of a butterfly, Neptis soma shania Evans 1924 (Lepidoptera: Nymphalidae) in the Sundarbans of Bangladesh. International Journal of Fauna and Biological Studies 1(3): 8-10.
Islam, Khandakar Kamrul and Rahman, Naimur. (2017). Two new records and one rediscovery of angiosperms for Bangladesh. Bangladesh J. Plant Taxon. 24(2): 227-231.
Islam, Khandakar Kamrul and Uddin, Sarder Nasir. (2021). Flemingia fluminalis C.B.Clarke (Fabaceae) - rediscovery of an angiosperm from Bangladesh. Bull. Bangladesh National Herb. 7: 149-151.
Khan, Md Kawsar and Hämäläinen, Matti. (2022). The rare and beautiful damselfly Rhinocypha trimaculata rediscovered in Bangladesh: With notes on earlier records, redescription of male and distinguishing characters from R. ignipennis (Odonata: Chlorocyphidae). Notulae odonatologicae 9(8): 315-330. https://doi.org/10.5281/zenodo.5702949
Rahman, M. Atiqur. (2017). Plant diversity inventory and assessment in the Hazarikhil Wildlife Sanctuary of Bangladesh and their conservation management plan. J. biodivers. conserv. bioresour. Manag. (under publ.) Aglaia edulis, Knema erratica, Artabotrys caudatus, Miliosa globosa]
Rahman, M. Atiqur and Rashid, M. Enamur. (2013). Status of endemic plants of Bangladesh and conservation management strategies. International Journal of Environment 2(1): 231-249. [Boesenbergia islamii]
Saha, Anik et al. (2022). A re-discovery of Coelops frithii (Chiroptera, Hipposideridae) from its type locality after one and a half century. Mammalia. https://doi.org/10.1515/mammalia-2021-0092
Shetu, Shayla Sharmin et al. (2022). An inventory of vascular flora of the Lalmai Hill area of Cumilla District, Bangladesh. Bangladesh J. Plant Taxon. 29(2): 203-240. https://doi.org/10.3329/bjpt.v29i2.63527
Sourav, Md Sharif Hossain, Anwar, Kabir Bin and Sultana, Mahbuba. (2020). Rediscovery of Dendrobium angulatum Lindl. (Orchidaceae), from Bangladesh after 198 years. Tropical Plant Research 7(2): 326-330.
Sourav, Md Sharif Hossain and Halder, Ronald. (2016). Rediscovery of Amorphophallus margaritifer (Roxb.) Kunth (Araceae) from Bangladesh. Tropical Plant Research 3(2): 361-363.
Uddin, Mohammad Zashim et al. (2011). Rediscovery of Osbeckia truncata D. Don ex Wight & Arn. (Melastomaceae) in Bangladesh. Bangladesh Journal of Botany 40(1): 101-103.
Chaetornis striata (Bristled grassbird)
Pelecanus philippensis (spot-billed pelican or grey pelican) [just a vagrant individual?]
Belarus
Belgium
DEKONINCK W. & GROOTAERT P., 2005. - Rediscovery of Lasius bicornis (FÖRSTER, 1850) in Belgium (Hymenoptera : Formicidae). Bulletin S.R.B.E./K.B.V.E., 141: 27–29.
Dekoninck, W. et al. (2004). Comments on rare and doubtful Belgian ant species and rediscovery of Myrmica lobicornis Nylander, 1846 (Formicidae, Hymenoptera). Bulletin S.R.B.E./K.B.V.E. 140: 31-33.
Dekoninck, W., Versteirt, V. and Grootaert, P. (2001). Rediscovery of a colony Polyergus rufescens (Latreille, 1798) in Belgium: Observations at the "Hageven" Nature reserve (Hymenoptera Formicidae). Bull. Soc. R. Belge Entomol. 137: 98-101. [Abstract]
Jacobs, Annelies and Jacobs, Steven. (2019). Rediscovery of Polycnemum majus (Amaranthaceae) in Belgium due to a 152 year old herbarium specimen. Dumortiera 114: 3-7.
Lock, Koen and Drumont, Alain. (2017). Rediscovery after seven decades of Limnephilus ignavus McLachlan, 1865 in Brussels and list of the caddisflies recorded for the botanical garden Jean Massart (Trichoptera: Limnephilidae). Bulletin de la Société royale belge d’Entomologie/Bulletin van de Koninklijke Belgische Vereniging voor Entomologie 153: 94-96.
Pollet, Marc et al. (2017). The rediscovery of Orthoceratium lacustre (Scopoli, 1763) (Diptera: Dolichopodidae) in Belgium, with data on its ecology and distribution in the Palaearctic region. Biologia 72(1): 62-69. [Abstract]
Sanabria, M. J., Lock, K., Scheepens, M. and Tempelman, D. (2012). Oligostomis reticulata (Linnaeus, 1761) (Trichoptera, Phryganeidae) recorded again in The Netherlands and Belgium. Lauterbornia 75: 1-13.
Sérusiaux, Emmanuël, Diederich, Paul, Ertz, Damien and van der Boom, Pieter. (2003). New or interesting lichens and lichenicolous fungi from Belgium, Luxembourg and northern France. IX (suite 1). Lejeunia, Revue de Botanique 173: [article 2].
Thomaes, Arno, Crevecouer, Luc and Wijnants, Michael. (2015). Tree cavity beetles in Haspengouw and Pays De Herve: Crepidophorus mutilatus (Elateridae) new for the Belgian fauna and rediscovery of Gnorimus variabilis (Cetoniidae) (Insecta: Coleoptera). Bulletin de la Société royale belge d’Entomologie/Bulletin van de Koninklijke Belgische Vereniging voor Entomologie 151: 40-51.
Vankerkhoven, François, Vanstraelen, Z. and Dekoninckd, W. (2011). Rediscovery of the parasitic ant Myrmica karavajevi (Arnoldi, 1930) in Belgium (Formicidae, Hymenoptera). Bulletin S.R.B.E./K.B.V.E. 147: 185-187.
Vanschoenwinkel, Bram, Brendonck, Luc, Pinceel, Tom, Dupriez, Pascal and Waterkeyn, Aline. (2013). Rediscovery of Branchipus schaefferi (Branchiopoda: Anostraca) in Belgium – notes on habitat requirements and conservation management. Belgian Journal of Zoology 143(1): 3-14.
VERSTEIRT V., DEKONINCK W., DESENDER K. & GROOTAERT P., 2002. - Rediscovery of Amara strenua in the ‘Uitkerkse polder’ area (Flanders, Belgium). Bulletin S.R.B.E./K.B.V.E., 138: 49 -52.
Belize
Balick MJ, Atha DE, Canham S, Romero L. (2002). Capoche - rediscovery of a forgotten febrifuge (Ocotea veraguensis [Meisn.] Mez) from Belize, Central America, including a new floristic record. Econ. Bot. 56(1): 89-91.
Bolivia
Delgado, Eliseo, Pacheco, Luis Fernando, Salazar-Bravo, Jorge and Rocha, Omar. (2018). Rediscovery of the chinchilla in Bolivia. Oryx 52(1): 13-15. [Abstract]
Gonzales, L., Lötters, S. and Reichle, S. (1999). On the dendrobatid frogs from Bolivia: rediscovery of Epipedobates bolivianus (Boulenger, 1902), first record of Colostethus brunneus (Cope, 1887) and comments on other species (Anura: Dendrobatidae). Herpetozoa 12: 179-186. [rediscovery of the Pilon Lajas Biosphere Reserve population after c.100 years]
Quinteros-Muñoz, Oliver et al. (2022). Signs of life: rediscovery of Erythrolamprus andinus (Dixon, 1983) (Serpentes: Dipsadidae) in Bolivia. Graellsia 78(1): e166. https://doi.org/10.3989/graellsia.2022.v78.341
Arachis ipaensis
Bosnia and Herzegovina
Koren, Toni and Martinović, Matea. (2019). The rediscovery of the Southern Swallowtail (Papilio alexanor Esper, 1799) in Bosnia & Herzegovina after 50 years of absence. Entomologist’s Gazette 70: 27-30.
Milanović, Đorđije. (2012). Liparis loeselii (L.) Rich. – a plant rediscovered in the Balkan peninsula. Botanica Serbica 36(2): 85-89.
Brazil
Alves, S.S., Bolzan, A.M.R., Santos, T.G., Gressler, D.T., Cechin, S.Z. (2013). Rediscovery, distribution extension and defensive behavior of Xenodon histricus (Squamata: Serpentes) in the state of Rio Grande do Sul, Brazil. Salamandra 49(4): 219-222.
Barbosa, J. L. et al. (2016). Rediscovery and range extension of the black-shouldered opossum Caluromysiops irrupta Sanborn, 1951 (Didelphimorphia, Didelphidae) in Brazil. Mammalia 80(3): 325-328. [Abstract]
Bencke, Glayson A. et al. (2018). Recent records of the Star-throated Antwren, Rhopias gularis (Spix, 1825) (Aves, Thamnophilidae), in Rio Grande do Sul, southern Brazil. Check List 14(6): 1099-1104.
Carvalho dos Reis, Luciana; Cassiano de Oliveira, Hermeson; Cavalcanti Pôrto, Kátia. (2020). Drepanolejeunea pinnatiloba Schiffn. (Lejeuneaceae): a rare species in Brazil rediscovered after 17 years. CheckList 16(4): 593-596.
Corrêa-do-Nascimento, Guilherme Sanches et al. (2020). The rediscovery of Rhodnius domesticus Neiva & Pinto, 1923 (Hemiptera: Reduviidae: Triatominae) in the state of Espírito Santo, Brazil. Rev Soc Bras Med Trop. 54:e03232020. doi: 10.1590/0037-8682-0323-2020.
Correa L. L. C.; D. E. Silva; L. H. Cappellari. (2010). Aves, Tinamidae, Crypturellus noctivagus noctivagus (Wied, 1820): Southward range extension and rediscovery in Rio Grande do Sul, Brazil. Check List 6(4): 585-586.
Costa, M., Muller, D. A., Saccol, S. S. A. and Santos, T. G. (2025). Rediscovery, distribution extension, and geographic distribution map of the Uruguay Harlequin Frog, Pseudis limellum (Cope, 1862) (Anura, Hylidae), in the Brazilian Pampa. Check List 22(2): 237-241. https://doi.org/10.15560/22.2.237
Cunha, Carlo M. and Migotto, Alvaro E. (2015). Rediscovery of Gastropteron chacmol (Gastropoda: Gastropteridae) on the Brazilian coast. Marine Biodiversity. doi:10.1007/s12526-015-0312-1 [Abstract]
de Oliveira, Arthur Schramm, de Souza, Wellington Vasconcelos, Schossler, Martin, Adams, Guilherme Bard and Balestrin, Rafael Lucchesi. (2026). Rediscovering the lizard Urostrophus undulatus (Squamata, Iguanidae) (Wiegmann, 1834) in southern Brazil. Boletín de la Sociedad Zoológica del Uruguay 35(1): e35.1.1. https://doi.org/10.26462/35.1.1
Filgueiras, T. S. and Shirasuna, R. T. (2009). Redescoberta de espécies presumivelmente extintas de Poaceae da Flora de São Paulo, Brasil. Hoehnea 36: 507-509. [Gymnopogon burchellii, Ichnanthus bambusiflorus, Leersia ligularis, Merostachys neesii, Paspalum falcatum]
Freitas, A. V. L., L. A. Kaminski, C. A. Iserhard, E. P. Barbosa & O. J. Marini Filho. 2011. The endangered butterfly Charonias theano (Boisduval) (Lepidoptera: Pieridae): current status, threats and its rediscovery in the state of São Paulo, southeastern Brazil. Neotrop. Entomol. 40(6): 669–676.
Gissi, Danilo Soares and Groppo, Milton. (2017). Parasites on the spot: the rediscovery of a presumably extinct Apodanthaceae in São Paulo state, Brazil. Check List 13(1): 2028. [Pilostyles blanchetti]
Gonçalves, Mateus Tomás Anselmo et al. (2022). Rediscovering Andreaea subulata Harv., a critically endangered moss in Brazil. Acta Botanica Brasilica 36: e20220165. https://dx.doi.org/10.1590/1677-941X-ABB-2022-0165
Just, João Paulo Gava, Colvero, Ricardo Dossa and Zocche, Jairo José. (2017). Rediscovery of Sporophila frontalis (Verreaux, 1869) (Aves, Thraupidae) in the state of Rio Grande do Sul, southern Brazil. Check List 13(2): 2069.
Kindel, Ivo and Kohler, Glauco. (2017). Rediscovery of Blackfronted Piping Guan Aburria jacutinga in the Itajaí Valley, Santa Catarina, southern Brazil. Cotinga 39: 50-51.
Macedo, J. M. et al. (2023). Rediscovery of Uzelothrips scabrosus Hood, 1952, a recent representative of a basal Thysanoptera (Arthropoda: Insecta) lineage, in Brazil. Brazilian Journal of Biology 83: e251434.
Maciel, A.O.; H.C. Costa; L.O. Drummond; J.O. Gomes; A. D’Angiolella. (2013). Rediscovery of Siphonops annulatus (Mikan, 1820) (Amphibia: Gymnophiona: Siphonopidae) in the state of Pará, Brazil, with an updated geographic distribution map, and notes on size and variation. Check List 9(1): 106-110.
Marinho-Filho, Jader and Veríssimo, Ed Wilson. (1997). The rediscovery of Callicebus personatus barbarabrownae in northeastern Brazil with a new western limit for its distribution. Primates 38(4): 429-433. [Abstract]
Marini, M. A., Duraes, R., Lopes, L. E., Fernandes, A. M. and de Alencar Carvalho, C. E. (2003). Rediscovery of Scalloped Antbird Myrmeciza ruficauda in Minas Gerais, Brazil. Cotinga 19: 59-61.
Marmontel, M., Calvimontes U., J. and Carvalho Jr, O. (2015). Rediscovery of Pteronura brasiliensis in the Amanã Sustainable Development Reserve, Amazonas, Brazil. Latin American Journal of Aquatic Mammals 10(2): 147-151.
Meller, Dante Andres, Frick, Ademir et al. (2021). The rediscovery of the Crested Eagle (Morphnus guianensis) in Rio Grande do Sul state, Brazil. Ornithology Research. https://doi.org/10.1007/s43388-021-00056-z [Abstract]
Meller, Dante Andres and Guadagnin, Demétrio Luís. (2016). Rediscovery of the Harpy eagle Harpia harpyja (Accipitriformes: Accipitridae) for Rio Grande Do Sul State, Brazil. Revista Brasileira de Ornitologia 24(1): 53-57.
de Melo, Alyson Vieira et al. (2021). Rediscovery and historical records of the Strange-tailed Tyrant Alectrurus risora (Passeriformes: Tyrannidae) in Brazil. Ornithology Research 29: 56-60. [Abstract]
Mendonça-Lima A, Zilio F, Joenck CM, Barcellos A (2006) Novos registros de Spizaetus ornatus (Accipitridae) no sul do Brasil. Revista Brasileira de Ornitologia 14(3): 279-282.
Parentes, Jaíne Maria Silva and Cavalcanti, Laise de Holanda. (2022). Rediscovery of Hemitrichia leiocarpa (Trichiales, Myxomycetes) in Brazil. Rodriguésia 73: e02242020. https://doi.org/10.1590/2175-7860202273042
de Pontes, Ricardo A. and Versieux, Leonardo M. (2018). Rediscovering the rare bromeliad Araeococcus goeldianus L.B. Smith (Bromelioideae) in Brazil and an amendment to its description. Phytotaxa 367(2): 186-190. [Abstract]
Roberto, I. J.; S. C. Ribeiro; M. M. S. Delfino; W. O. Almeida. (2009). Reptilia, Colubridae, Helicops angulatus: distribution extension and rediscovery in the state of Ceará. Check List 5(1): 118-121.
Sagot-Martin, François et al. (2020). An updated checklist of the birds of Rio Grande do Norte, Brazil, with comments on new, rare, and unconfrmed species. Bull. B.O.C. 2020 140(3): 218-298. [Crypturellus noctivagus (ethnoknown) and Procnias averano]
Sales, Licia, Queiroz, Vinicius and Migotto, Alvaro E. (2016). Rediscovery of Okenia impexa Marcus 1957 on the Brazilian coast: the first image of a living specimen from Brazil. Spixiana 39(2): 203-204.
Santos, Diego, Costa, Swami Leitão and Buril, Maria Teresa. (2022). Nomenclatural, taxonomic, and distribution novelties concerning Evolvulus chrysotrichos (Convolvulaceae): a rare and endangered species from Brazil. Rodriguésia 73: e01452021. https://doi.org/10.1590/2175-7860202273081
Santos, T. G.; C. F. B. Haddad. (2006). Amphibia, Anura, Leptodactylidae, Leptodactylus labyrinthicus: rediscovery and distribution extension in the state of Rio Grande do Sul, Brazil. Check List 2(1): 22-23.
Siewert, Ricardo Russo et al. (2016). Stichelia pelotensis (Lepidoptera, Riodinidae): conservation, notes, and rediscovery of an endangered butterfly from southern Brazil. Revista Brasileira de Entomologia 60: 105-108.
Silva, Eduardo Oliveira, Milward-de-Azevedo MA, Ferreira AWC, Sobral MEG (2020) Rediscovery and new records of Passiflora auriculata Kunth and P. cisnana Harms (Passifloraceae) in Brazil. Check List 16(2): 441-449.
Simon, José Eduardo and Magnago, Gustavo rodrigues. (2013). Rediscovery of the cryptic Forest-Falcon Micrastur mintoni Whittaker, 2002 (Falconidae) in the atlantic forest of southeastern Brazil. Revista Brasileira de Ornitologia 21(4): 257-262.
Spielmann, Adriano Afonso et al. (2021). The rediscovery of Xanthoria (Teloschistaceae) in Brazil. Archive For Lichenology 24: 1-7.
Tomas, Walfrido Moraes et al. (2015). The Carandilla Palm (Trithrinax schizophylla Drude, Arecaceae) is not extinct in Brazil: first primary records from the Chaco region of Mato Grosso do Sul. Check List 11(4): 1669.
João Filipe Riva Tonini, Israel Souza Mendonça, Alyson Borges Coutinho and João Luiz Gasparini. (2011). Anurans from Costa Bela, state of Espírito Santo, southeastern Brazil: inventory at an urban area and the re-discovery of Allobates in the state. Herpetology Notes 4: 435-444. [Allobates olfersioides]
Velloso, Jorge Renato Pinheiro, Putzke, Jair and Cavalcanti, Laise de Holanda. (2022). Lycogala flavofuscum (Liceales, Myxomycetes): a rare species in South America, rediscovered in Brazil after 100 years. Rodriguésia 73: e01442021. https://doi.org/10.1590/2175-7860202273080
Zank, C.; I. L. Kaefer; P. Colombo; R. Lingnau; A. P. Santos-Jr; C. Both; F. M. D'Agostini; R. C. Santos; S. Z. Cechin. (2008). Amphibia, Anura, Leptodactylidae, Leptodactylus furnarius: Rediscovery and distribution extension in the state of Rio Grande do Sul, Brazil. Check List 4(1): 89-91.
"rediscovery of Morpho epistrophus nikolajewna (Morphini) in a highly urbanized forest fragment in northeast Brazil."
Britain
Alexander, K. N. A. (2007). Lebia chlorocephala (Hoffmannsegg) (Carabidae) rediscovered in Devon after 100 years. The Coleopterist 16(3): 124.
Allan., P.B.M., 1965 The New Forest Cicada. The Entomologist's Record and Journal of Variation 77: 295-296.
Balfour-Browne, F. (1909). Notes on the rediscovery of Apus cancriformis in Britain. Annales of Scottish Natural History 1909: 118.
Balfour-Browne F. 1948. Re-discovery of Apus cancriformis. Nature 162:116
Beaumont, H. E. (2019). Nematopogon magna (Zeller, 1878) (Lep.:Adelidae). Rediscovery inYorkshire after more than 100 years. The Entomologist’s Record and Journal of Variation 131: 150-151.
Davy-Bowker, John et al. (2018). Rediscovery of the critically endangered ‘scarce yellow sally stonefly’ Isogenus nubecula in United Kingdom after a 22 year period of absence. Zootaxa 4394(2): 295-300. [Abstract]
Efford, Ian. (1959). Rediscovery of Bathynella chappuisi Delachaux in Britain. Nature 184: 558-559.
Fay, M. F. (2013). 751. ORCHIS MILITARIS. Curtis's Botanical Magazine 30: 9-17.
Foster, C. W. and Holloway, G. J. (2018). Dicranocephalus agilis (Scopoli) (Hemiptera, Heteroptera) at Slapton Sands. Field Studies [2018]: 4 pp.
Fox HM. 1949. On Apus: its rediscovery in Britain, nomenclature and habits. Proceedings of the Zoological Society of London 119:693-702
Gerlach, Justin. (2014). Extinct Animals of the British Isles. Phelsuma Press. 96 pp. [Lithophane furcifera furcifera]
Golding, N., Nunn, M. A., Medlock, J. M., Purse, B. V., Vaux, A. G. C. and Schafer, S. M. (2012). West Nile virus vector Culex modestus established in southern England. Parasites & Vectors 5(1): 32.
Long, David G. (2021). Grimmia anodon rediscovered on Arthur's Seat. Field Bryology 88: 2-3.
McCulloch, James I. (2021). Amphimallon fallenii (Gyllenhal) (Scarabaeidae: Melolonthinae) rediscovered in south-west England. The Coleopterist 30(2): 94-100.
Rackley, Adele (9 February 2012). Disease-spreading mosquito found in UK after 60 years. Planet Earth Online. UK Natural Environment Research Council.
Russell-Smith A (2017) The rediscovery of Nigma flavescens in Britain. Newsletter of the British Arachnological Society 140: 16-17.
Turner, Clive R. (2008). On the rediscovery of Lebia chlorocephala (Hoffmann, J.) (Carabidae) in Devon. The Coleopterist 17(2): 99-100.
Walters, S. M. (1974). The rediscovery of Senecio paludosus L. in Britain. Watsonia 10(1): 49-54. [now Jacobaea paludosa angustifolius]
Williams P, Whitfield M, Biggs J, Bray S, Fox G, Nicolet P, Sear D. 2004. Comparative biodiversity of rivers, streams, ditches and ponds in an agricultural landscape in Southern England. Biological Conservation 115(2): 329-341. [Abstract] [Triops cancriformis] [source: Sellers GS, Griffin LR, Hänfling B, Gómez A. (2017) A new molecular diagnostic tool for surveying and monitoring Triops cancriformis populations. PeerJ 5:e3228]
Wolton, Robert J. and Drake, C. Martin. (2015). Rhaphium pectinatum (Loew) (Diptera, Dolichopodidae) re-found in Britain. Dipterists Digest 22: 1-4.
Catocala fraxini (Clifden nonpareil)
Greater mouse-eared bat (Myotis myotis)
Cambridgeshire
Anon. (1976). Fen ragwort rediscovered. Conserv. Rev., no. 13. 8.[Senecio paludosus]
Hampshire
Khalaf, A. N. (1978). Some notes on the biology and rediscovery of the tadpole shrimp, Triops (Apus) cancriformis Bosc. (Notostraca: Branchiopoda) in Hampshire, Britain. Bulletin of the Natural History Research Centre (Iraq) 7(2): 29-34.
Herefordshire
Villa cingulata (Downland villa)
Isle of Wight
Beavan, S. D. (2021). Dialectica imperialella (Zeller, 1847) (Lepidoptera: Gracillariidae) rediscovered on the Isle of Wight after 45 years and consideration of one of its larval foodplants. Entomologist's Gazette 72(4): 221-228. https://doi.org/10.31184/G00138894.724.1835
Pope, Colin. (2017). The Marshlands of Freshwater, a case study. Proc. Isle of Wight Nat. Hist. Archaeol. Soc. 31: 37-57. [Valeriana dioica, the Marsh valerian]
Sussex
Scarce Hook-tip (Sabra harpagula)
Bulgaria
Assyov, Boris, Bozok, Fuat, Slavova, Monica and Taşkın, Hatıra. (2023). Rediscovery and ITS-barcoding of Entonaema cinnabarinum
(Xylariales, Ascomycota) from Bulgaria. Historia Naturalis Bulgarica 45(8): 197-210. https://doi.org/10.48027/hnb.45.081
Assyov, Boris and Dimitrov, Dimitar. (2022). Pluteus fenzlii (Pluteaceae, Agaricales) Rediscovered in the Balkan Peninsula after over 150 Years Gap. Ecologia Balkanica 14(1): 155-160.
Assyov, Boris and Petrova, A. (eds.). (2012). Conspectus of the Vascular Flora of Bulgaria. Distribution maps and floristic elements. BBF, Fourth revised and enlarged edition, S. 490 p. (in Bulgarian). [Hammarbya paludosa and Liparis loeselii]
Bekchiev, Rostislav et al. (In Press, 2020). One Hundred Years of Solitude: Rediscovery of the Rare and Protected Beetle Rhysodes sulcatus (Fabricius, 1787) (Coleoptera: Rhysodidae) in Bulgaria. Acta Zool. Bulg.
Bekchiev, Rostislav et al. (2022). Carabus variolosus (Fabricius, 1787) (Coleoptera: Carabidae) in Bulgaria: rediscovered after 111 years. Historia Naturalis Bulgarica 44(10): 119-123.
Beshkov, S. 2014. Field Guide to the Butterflies and Skippers (Papilionoidea and Hesperioidea) from Vitosha Nature Park, Bulgaria. II. Directorate of the Vitosha Nature Park, Sofia. 222 p. (In Bulgarian). [Nymphalis xanthomelas]
Hoejgaard, K. and Beshkov, S. (2011). Rediscovering Muschampia tessellum (Hübner, [1803]) (Lep.: Hesperiidae) in Bulgaria with Additional Notes on M. cribrellum (Eversmann, 1841) from the Eastern Balkan (Stara Planina) Mountains. – The Entomologist’s Record and Journal of Variation 123: 147-150.
Marinov, Y. et al. (2014). Current status of the rare species Lathyrus transsilvanicus in Bulgaria. Bulgarian Journal of Agricultural Science 20 (Supplement 1): 160-164.
Cambodia
Goes, F., Hong C., Suon M., Luon K. and Meas, R. (1998). Waterbird counting and survey at Prek Toal and Boeng Chhma/ Moat Khla. May – June 1998. Tonle Sap Technical Coordination Unit, Ministry of Environment, Phnom Penh, Cambodia. [Heliopais personata]
Platt, S.G., Stuart, B.L., Sovannara, H., Kheng, L., Kalyar & Kimchhay, H. (2003) Rediscovery of the critically endangered river terrapin, Batagur baska, in Cambodia, with notes on occurrence, reproduction and conservation status. Chelonian Conservation and Biology 4: 691-695.
Hog deer: http://www.phnompenhpost.com/national/hello-deer
Cameroon
Onana, Jean Michel, Cheek, Martin and Pollard, Benedict John. (2011). Red Data Book of the Flowering Plants of Cameroon: IUCN Global Assessments. Kew, Richmond, Surrey, UK: Kew Publishing. 578 pp. [Sclerochiton preussii (p. 74); Trichostachys interrupta (p. 308); Habenaria microceras (p. 402)]
Canada
Bleakney, J. S. and Bailey, K. H. (1967). Rediscovery of the salt-marsh sacoglossan Alderia modesta Loven in eastern Canada. Journal of Molluscan Studies, 37(5), 347-349.
Picard, Isabelle et al. (2009). Modern Records of the Pink Heelsplitter Mussel, Potamilus alatus (Say, 1817), in the Ottawa RiverDrainage, Québec and Ontario, Canada. Northeastern Naturalist 16(3): 355-364.
Short-flower suncup (Taraxia breviflora)
Taylor’s checkerspot butterfly
British Columbia
Kenner, Rex D. and Cannings, Robert A. (2000). Rediscovery of the Western Pondhawk in the Lower Mainland. Discovery [2000](Winter): 50-52.
Kimoto, Troy and Buck, Matthias. (2015). Rediscovery of Cerceris fumipennis (Hymenoptera: Crabronidae) in British Columbia, Canada, with notes on geographic variation and nesting habits. The Canadian Entomologist 147(4): 419-424.
Ontario
Lewis, Christopher J. (2019). Heterodermia leucomela (L.) Poelt discovered in Ontario, Canada for the first time in over 150 Years. Evansia 36(2): 30-38.
Canary Islands
Verloove, F., Sennikov, A. N., Reyes-Betancort, J. A. (2023). Taxonomy and nomenclature of Abutilon albidum (Malvaceae, Malvoideae), a cryptic Saharo-Canarian species recently rediscovered in Tenerife. PhytoKeys 221: 41-60. https://doi.org/10.3897/phytokeys.221.95907
Chile
Araya, Juan Francisco and Bitner, Maria Aleksandra. (2018). Rediscovery of Terebratulina austroamericana Zezina, 1981 (Brachiopoda: Cancellothyrididae) from off northern Chile. Zootaxa 4407(3): 443-446. [Abstract]
Lamilla-Maulén, Pablo et al. (2026). The Largest Nematogenys inermis From a Small Stream in the Maipo Basin Once Considered Extinct: Perspectives on Body Size Evolution and Peripheral Habitats as Refuges. Austral Ecology 51(3): e70202. https://doi.org/10.1111/aec.70202
China
Arai, R., Xie, Y. and Akai, Y. (1995). Rediscovery of the bitterling, Tanakia lanceolata, in China (Pisces, Cyprinidae). Japanese Journal of Ichthyology 42(2): 196-199.
Byun, Bong-Kyu and Yan, Shanchun. (2005). Description of a new species, records of five previously unrecorded species, and rediscovery of a lost species in the genus Ancylis Hübner (Lepidoptera: Tortricidae) from China. Zootaxa 1103: 17-26. [Abstract]
Chen, Zhi-Teng and Du, Si-Kai. (2020). Rediscovery of the only known salmonfly from China, Pteronarcys sachalina Klapálek, 1908 (Plecoptera: Pteronarcyidae). Aquatic Insects 41(2): 174-183. [Abstract]
Du, H.; Li, L.; Wei, Q.; Zhang, S.; Wang, C.; Sun, Q.; Yang, X.; Li, L., 2014: The rediscovery of Hucho bleekeri in the Taibai River, the upper tributary of the Hanjiang River, China. Chin. J. Zool. 3, 414 (In Chinese).
Fei Li, Xi Zheng, Xue-Long Jiang and Bosco Pui Lok Chan. (2017). Rediscovery of the sun bear Helarctos malayanus in Yingjiang County, Yunnan Province, China. Zoological Research 38(4): 1-11. [automatic download]
Guo, Wei et al. (2017). The rediscovery of Large-spotted Civet Viverra megaspila in China. Small Carnivore Conservation 55: 88-90.
Jiang, Kai-Wen et al. (2022). Legume additions to the flora of China. Phytotaxa 532(1): 1-21. https://doi.org/10.11646/phytotaxa.532.1.1 [Flemingia vestita]
Liang, Tao et al. (2021). Rediscovery of Ablepharus deserti (Squamata: Scincidae) from the northwest of China, including a northeastward range extension for this species. Herpetology Notes 14: 1373-1374.
Wang, Ji-Shen. (2022). New and little-known species of the genus Dicerapanorpa from northwestern Yunnan, China (Mecoptera: Panorpidae). Acta Entomologica 62(1): 1-13. [Dicerapanorpa triclada]
Xia, X.-R., Zheng, J.-Y., Zhang, D.-N., Zhang, S.-B., Wu, Y.-H. and Huang, W.-Z. (2025). Notes on Meesiaceae (Splachnales) in China: confirmation of Meesia minor, and rediscovery of Amblyodon dealbatus after nearly thirty years. Herzogia 38(2): 262-274. https://doi.org/10.13158/heia.38.2.2025.262
China (Hong Kong)
Yan, Yue-Hong et al. (2004). Rediscovered ferns from Hong Kong, China. Porcupine 31: 12-13. [Trichomanes auriculatum, Hypolepis punctata, Cyrtomidictyum basipinnatum, Chieniopteris kempii]
Zhejiang
Mengjun, Sun, Zhenghai, Chen, Weisong, Weng, Shundong, Ying and Qun, Du. (2001). New records on the distribution of the rare and endangered plants in Zhejiang. Journal of Zhejiang Forestry Science and Technology 21(4): 7-10. [Abstract] [Ranalisma rostratum, Disanthus cercidifolius longipes]
Colombia
Bryce, G.W., Jr. 1970 Rediscovery of the limpet, Acroloxus colorandensis (Basommatophora: Acroloxidae), in Colorado. Nautilus 83: 105-108.
Colorado, Gabriel J. (2008). Rediscovery of the Recurve-billed Bushbird for the Cordillera Central of Colombia. Ornitologia Neotropical 19: 467-471.
Fernández-Roldán, Juan David, Lynch, John D. and Bocanumenth, Daniel. (2022). Rediscovery of Oscaecilia ocrocephala (Cope, 1886) (Amphibia: Gymnophiona: Caeciliidae) in Colombia, with comments on its variation and affinity to O. polyzona (Fischer, 1880). Revista Latinoamericana de Herpetología 5(3): 25-31. https://doi.org/10.22201/fc.25942158e.2022.3.453
Kattan, Gustavo H. and Beltrán, J. William. (1997). Rediscovery and status of the Brown-banded Antpitta Grallaria milleri in the central Andes of Colombia. Bird Conservation International 7: 367-371.
Mendieta-Giraldo, L., Escobar-Lasso, S., Grajales-Suaza, E. and González-Maya, J.F. (2021). Not all gone: the rediscovery of Jaguar (Carnivora: Felidae: Panthera onca) and records of threatened monkeys (Primates: Mammalia) in the Magdalena River Valley of Caldas Department in Colombia, a call for their conservation. Journal of Threatened Taxa 13(3): 17865-17874. https://doi.org/10.11609/jott.6673.13.3.17865-17874
Costa Rica
Angulo, Arturo et al. (2022). Rediscovery of the endangered Amphilophus lyonsi (Gosse 1966) (Cichliformes: Cichlidae) in Costa Rica; extended diagnosis and description and new ecological data. Zootaxa 5182(6): 528-540. https://doi.org/10.11646/zootaxa.5182.6.2
Chaves, G., Zumbado-Ulate, H., García-Rodríguez, A., Gómez, E., Vredenburg, V. T. and Ryan, M. J. (2014). Rediscovery of the Critically Endangered streamside frog, Craugastor taurus (Craugastoridae), in Costa Rica. Tropical Conservation Science 7(4): 628-638.
Kubicki, B. (2004). Rediscovery of Hyalinobatrachium chirripoi (Anura: Centrolenidae) in southeastern Costa Rica. Revista de Biología Tropical 52: 215-218.
Pineda, W., Rodríguez-Herrera, B. and Timm, R. M. (2008). Rediscovery, ecology, and identification of rare free-tailed bats (Chiroptera: Molossidae) in Costa Rica. Acta Chiropterologica 10(1): 97-102.
Croatia
Bogdanović, Sandro, Šegota, Vedran and Alegro, Antun. (Accepted, 2018). Resurrection of a regionally extinct taxon in Croatia – the case of Ammophila arenaria (L.) Link (Poaceae). Acta Botanica Croatica. DOI: 10.2478/botcro-2018-0013
Gljušćić, Edi, Bilajac, Andrea and Iveša, Ljiljana. (2022). Rediscovery of Digenea simplex (Wulfen) C. Agardh 1822 (Rhodomelaceae, Ceramiales) in the northern Adriatic, p. 159. In: University of Zagreb, Faculty of Science, 6th PhD Student Symposium 2022, April 23-24, 2022.
Justić, M., Bučar, M., Vizec, P., Vukres, A., Šegota V. & Vuković, N. (2020). The rare and endangered orchid Cypripedium calceolus L. in Croatia - refound in Gorski Kotar (West Croatia) after 126 years. Nat. Croat. 29(1): 55-62.
Mihoci, Iva et al. (2012). Rediscovery of the threatened butterfly Nymphalis vaualbum in Croatia with remarks on its historical findings. Nartura Croatica 21(1): 259-262.
Podnar, M., Mauch Lenardić, J., Kulijer, D., Ljubas, V., Hamidović, D. and Tvrtković, N. (2024). Rediscovery of the blind mole rat in Croatia with revision of the phylogenetic analysis of the Nannospalax
monticola complex in Europe. Nat. Croat. 33(2): 317-336.
Šegota, Vedran, Hršak, Vladimir and Alegro, Antun. (2016). Long time no see – rediscovery of peculiar ephemeral fern Anogramma leptophylla (L.) Link in Croatia. Acta Botanica Croatica 76(1): 91-94.
Skejo, Josip et al. (2023). Endangered Transsylvanian wingless groundhopper (Tetrix transsylvanica) is not extinct in Croatia and requires urgent protection. Nat. Croat. 32(1): 241-255. [Tetrix transsylvanica hypsocorypha]
Verovnik, R. and Švara, V. (2016). Confirmed recent occurrence of the Southern Swallowtail (Papilio alexanor Esper, 1799) in Croatia (Lepidoptera: Papilionidae). SHILAP Revta. lepid., 44(176): 547-552.
Cuba
Krings, A., Areces Berazaín, F. and Lazcano Lara, J. C. (2005). New and rediscovered milkweeds from Cuba: Calotropis gigantea and Gonolobus stephanotricus (Apocynaceae: Asclepiadoideae). Willdenowia 35: 315-318. [Gonolobus stephanotricus]
Cyprus
Aristophanous, Marios, Pittaway, Anthony R. and Aristophanous, Aristos. (2022). Rediscovery of Clarina syriaca (Lederer, 1855) (Lepidoptera, Sphingidae, Macroglossinae) in Cyprus after 70 years; with notes on its biology and early life history from the Levant. Nota Lepi. 45: 305-314.
Sparrow, David J. et al. (2020). First records of Aeshna isoceles and the rediscovery of Lestes barbarus on Cyprus (Odonata: Lestidae, Aeshnidae). Notulae odonatologicae 9(5): 185-195.
Czech Republic
Brabec, J. and Honsa, P. (2004). Rerisnicnik skalni (Cardaminopsis petraea) - znovunalezeny druh CHKO Krivoklatsko. (Cardaminopsis petraea - rediscovered species of the Krivoklatsko Biosphere Reserve (Central Bohemia).) Zpravy Ceske Bot. Spol. (Prague) 39. 387-392.
Chvojka, P., Komzák P. and Špaček J. (2009). New faunistic records of Trichoptera (Insecta) from the Czech Republic, III. Acta Musei Moraviae, Scientiae biologicae (Brno), 94: 81-85. [Hydroptila occulta]
Halačka, Karel et al. (2017). A newly discovered population of the Balkan spiny loach Sabanejewia balcanica (Karaman, 1922) in the River Jihlava, Czech Republic. Folia Zoologica 66(3): 163-166. [Abstract]
Jurajda, Pavel and Pavlov, Igor. (2016). Rediscovery of Rutilus virgo in the River Dyje, Czech Republic. Folia Zoologica 65(2): 98-100. [Abstract]
Komzák, Petr and Kroča, Jiří. (2011): New faunistic records of Trichoptera (Insecta) from the Czech Republic, IV. Acta Musei Moraviae, Scientiae Biologicae 96(1): 189-192. [Hydroptila tineoides]
Komzák, Petr & Kroča, Jiří. (2018). New faunistic records of Hydroptilidae (Insecta, Trichoptera) from the Czech Republic. Acta Mus. Siles. Sci. Natur., 67: 165-173. [Oxyethira tristella]
Malíček J., Berger F., Bouda F., Cezanne R., Eichler M., Halda J. P., Langbehn T., Palice Z., Šoun J., Uhlík P. & Vondrák J. (2017). Lišejníky zaznamenané během bryologicko-lichenologického setkání v Mohelně na Třebíčsku na jaře 2016 [Lichens recorded during the bryological and lichenological meeting in Mohelno (Třebíč region, southwestern Moravia) in spring 2016]. Bryonora 60: 24-45. [Arthopyrenia analepta]
Peterka, Tomáš et al. (2021). Drepanocladus lycopodioides rediscovered in the Czech Republic: Abandoned quarries as refugia for endangered fen species. Aquatic Botany 172: 103380. [Abstract]
Procházka, Jiří et al. (2020). Rediscovery of the rare saproxylic beetle Peltis grossa (Linnaeus, 1758), Coleoptera: Trogossitidae, in the Šumava National Park and its occurrence in surrounding area. Silva Gabreta 26: 33-49.
Vondrák, Jan et al. (2022). From Cinderella to Princess: an exceptional hotspot of lichen diversity in a long-inhabited central-European landscape. Preslia 94: 143-181. [Arthopyrenia fraxini, Callome multipartita, Dermatocarpon meiophyllizum, Ramalina obtusata, Rinodina archaea]
Denmark
Fischer, Frederik Leck, Prehn, Johan Kjær and Hansen, Aslak Kappel. (2026). New Danish Spider Records Confirmed through DNA Barcoding: Simziella major (Menge, 1869) and Synageles hilarulus (C. L. Koch, 1846). Ent. Medd. 92(1-2): 33-42. [Simziella major (a spider) after six decades]
Dominica, Commonwealth of
Ecuador
Balslev, H. (1979). On the distribution of Rostkovia magellanica (Juncaceae), a species newly rediscovered in Ecuador. Brittonia 31(2): 243 - 247.
Henning, Tilo, Acuña-Castillo, Rafael, Cornejo, Xavier, Gonzáles, Paúl, Segovia, Edgar, Wong Sato, Akira Armando and Weigend, Maximilian. (2023). When the absence of evidence is not the evidence of absence: Nasa (Loasaceae) rediscoveries from Peru and Ecuador, and the contribution of community science networks. PhytoKeys 229: 1-19. https://doi.org/10.3897/phytokeys.229.100082 [Nasa ramirezii]
Valdiviezo-Rivera, J., Buenaño Carriel, M. and Escobar-Camacho, D. (2021). Rediscovery of Hemigrammus unilineatus (Gill, 1858) (Characiformes, Characidae) in Ecuador after more than three decades. Check List 17(4): 1181-1185. https://doi.org/10.15560/17.4.1181
Egypt
García, N., Cuttelod, A. and Abdul Malak, D. (eds.) (2010). The Status and Distribution of Freshwater Biodiversity in Northern Africa. Gland, Switzerland, Cambridge, UK, and Malaga, Spain: IUCN. xiii+141pp. [Agriocnemis sania]
Serag, Mamdouh S. (2000). The Rediscovery of Papyrus (Cyperus papyrus L.), on the bank of the Damietta branch of the Nile Delta, Egypt. Taeckholmia 20(2): [pagination?]. [Abstract]
Ethiopia
Atkinson P., Robertson P., Dellelegn Y., Wondafrash M., Atkins J. (1996). The recent rediscovery of White-winged Flufftails in Ethiopia. African Bird Club 3(1): 34-36.
Fiji
Clause, Adam G., Thomas-Moko, Nunia, Rasalato, Sialisi and Fisher, Robert N. (2018). All is not lost: Herpetofaunal “extinctions” in the Fiji Islands. Pacific Science 72(3): [pagination?].
Morrison, C., Naikatini, A., Thomas, N., Rounds, I., Thaman, B and Niukuln, J. (2004). Rediscovery of an endangered frog Platymantis vitianus, on mainland Fiji: implications for conservation and management. Pacific Conservation Biology 10(4): 237-240.
Finland
Evertsen, J.; Bakken, T.; Green, S. Rediscovery of Tenellia adspersa (Nudibranchia) from the Finnish archipelago. Sarsia 2004, 89, 362–365.
Lehtonen, Samuli, Li, Zhi-Zhong and Chen, Jin-Ming. (2026). Herbariomics and the peculiar case of Alisma wahlenbergii (Alismataceae). Taxon 75(1): e70097. https://doi.org/10.1002/tax.70097
Vahtera, Varpu and Lehtinen, Pekka T. (2018). Rediscovery of Geophilus carpophagus Leach (Chilopoda: Geophilomorpha) from Finland. Memoranda Soc. Fauna Flora Fennica 94: 36-38. [automatic download]
France
Denux, Olivier et al. (2017). Rediscovery of the endangered species Harpalus flavescens (Coleoptera: Carabidae) in the Loire River. Insect Conservation and Diversity 10(6): 488-494. https://doi.org/10.1111/icad.12228 [Abstract]
Nienhuis, J. A. J. H. (2003). The rediscovery of Spengler's freshwater pearlmussel Pseudunio auricularius (Spengler, 1793) (Bivalvia, Unionoidea, Margaritiferidae) in two river systems in France, with an analysis of some factors causing its decline. Basteria 67: 67-86.
Pires, Mathias et al. (2021). Redécouverte de Trifolium isthmocarpum Brot. en France continentale, espèce non revue depuis plus de cent ans. Carnets botaniques 69: 1-4.
Corsica
Scalera, R. and Angelici, F. M. 2002. Rediscovery of the Apennine Hare Lepus corsicanusin Corsica. Museo Regionale di Scienze Naturali Bollettino (Turin) 20(1): 161-166.
French Guiana
Massary, J.C., Hoogmoed, M.S. & Blanc, M. (2000) Comments on the type specimen of Dracaena guianensis Daudin, 1801 (Reptilia: Sauria: Teiidae), and rediscovery of the species in French Guiana. Zoologische Mededelingen, Leiden 74: 167-180.
Scharf, Uwe, Maas, Paul J. M. and Morawetz, Wilfried. (2006). Guatteria richardii (Annonaceae) rediscovered along with two new species from French Guiana. Blumea - Biodiversity, Evolution and Biogeography of Plants 51(3): 541-552. https://doi.org/10.3767/000651906X622094
Georgia
Beridze, Tamar et al. (2021). Ship sturgeon rediscovered in the Rioni River in Georgia. Oryx 55(1): 9.
Tuniyev B.S., Iremashvili G.N., Petrova T.V. and Kravchenko M.A. 2018. Rediscovery of the steppe viper in Georgia. Proceedings of the Zoological Institute RAS, 322(2): 87–107. https://doi.org/10.31610/trudyzin/2018.322.2.87 [Abstract]
Germany
Balkenhol, Birgit et al. (2022). The Great Raft Spider Dolomedes plantarius rediscovered in Saxony (Araneae: Pisauridae). Arachnologische Mitteilungen / Arachnology Letters 63: 24-29.
Bauer, Tobias, Thorns, H.-J. and Guttenberger, J. (2022). Rediscovery of the extremely rare running crab spider Philodromus (Artanes) poecilus (Thorell, 1872) in Bavaria, Germany, after 141 years (Araneae, Philodromidae). Check List 18(3): 583-591. https://doi.org/10.15560/18.3.583
Cezanne, R. and Eichler, M. (2013). Neue Funde von Parmotrema reticulatum (Taylor) M. Choisy in Hessen und Rheinland-Pfalz. Hessische Floristische Briefe 6 (2): 17-24.
Krüger, Andreas and Tannich, Egbert. (2013). Rediscovery of Anopheles algeriensis Theob. (Diptera: Culicidae) in Germany after half a century. Journal of the European Mosquito Control Association 31: 14-16.
Kuhlisch, Cornelius, Kampen, Helge and Walther, Doreen. (2018). Rediscovery of Culex (Neoculex) martinii Medschid, 1930 (Diptera, Culicidae) in Germany. Parasitology Research 117(10): 3351-3354. [Abstract]
Liston, Andrew D. and Skacel, Peter. (2015). The reappearance of Pseudoclavellaria amerinae (Linnaeus, 1758) in Germany (Hymenoptera, Cimbicidae). Contributions to Entomology 65 (1): 125-129.
Mrácek, Z., Weiser, J., Bureš, M. and Kahounová, L. (1992). Steinernema kraussei (Steiner, 1923) (Nematoda – Steinernematidae) rediscovery of its type locality in Germany. Folia Parasitologica 39: 181-182.
Müller, J. (2020). Wiederbesiedlung des Bayerischen Waldes durch Peltis grossa (Linnaeus, 1758). Faunistische Notiz, Arbeitsgemeinschaft bayerischer Entomologen. abeentomofaunistik.org
Müller, Reinhard and Hendrich, Lars. (2022). Wiederfund von Stenelmis canaliculata (Gyllenhal, 1808) (Coleoptera: Elmidae) in Niedersachsen nach über 100 Jahren. Lauterbornia 88: 75-81.
Neiber, Marco T. (2017). 'Auf der Reeperbahn nachts um halb eins' — Wiederfund des Bierschnegels in Hamburg nach 80 Jahren. Mitt. dtsch. malakozool. Ges. 96: 1-6. [Limacus flavus]
Otte, V., van den Boom, P. & Rätzel, S. 2006. Bemerkenswerte Funde von Flechten und lichenicolen Pilzen aus Brandenburg XI. Verhandlungen des Botanischen Vereins von Berlin und Brandenburg 139: 275-291. [Parmotrema reticulatum]
Raabe, U. (2009). Chara baueri rediscovered in Germany—Plus additional notes on Gustav Heinrich Bauer (1794–1888) and his herbarium. Int. Res. Group Charophytes News 20: 13-15.
Rychła, Anna and Buczyński, Paweł. (2003). Wiederfund von Leucorrhinia caudalis in Sachsen (Odonata: Libellulidae) [Rediscovery of Leucorrhinia caudalis in Saxony (Odonata: Libellulidae)]. Libellula 22(3-4): 119-125.
Werner, D. (2003). The Simuliidae (Diptera) of the River Oder and its tributaries, with special reference to the reappearance of Simulium (Schoenbaueria) nigrum (Meigen) in larger rivers in Central Europe. Journal of Natural History 37: 1509-1528. https://doi.org/10.1080/00222930210129287
Ghana
Anonymous. (2004). White-necked Picathartes rediscovered. Bulletin of the African Bird Club 11(2): 95-96.
Brichieri-Colombi, T. A. et al. (2016). In aid of (re)discovered species: maximizing conservation insights from minimal data. Animal Conservation. DOI: 10.1111/acv.12306
East, Rod. (1990). Antelopes: global survey and regional action plan. Part 3: West and Central Africa. IUCN, Gland, Switzerland. [Tragelaphus spekii gratus]
Marks, B. D. et al. (2004). Rediscovery of the White-necked Picathartes Picathartes gymnocephalus in Ghana. Bull. Br. Orn. Club 124: 151-153.
Greece
Bogaerts, Sergé et al. (2018). Rediscovery of the grass snake (Natrix natrix) on the island of Karpathos, Greece. Herpetology Notes 11: 303-305.
Strid, Arne et al. (2017). Additions to the flora of the Prespa National Park, Greece. Phytologia Balcanica 23(2): 207-269. [Aldrovanda vesiculosa in Lake Mikri Prespa]
Yannitsaros, Artemios G. and Constantinidis, Theophanis A. (1996). The rediscovery of Biebersteinia orphanidis Boiss. (Geraniaceae) in Greece. Botanical Journal of the Linnean Society 120: 239-242.
Guatemala
Bosland, Paul W. and Gonzalez, Max M. (2000). The rediscovery of Capsicum lanceolatum (Solanaceae), and the importance of nature reserves in preserving cryptic biodiversity. Biodiversity & Conservation 9(10): 1391-1397. https://doi.org/10.1023/A:1008930931976
Gulf of Guinea
Ayissi, Isidore et al. (2014). Rediscovery of Cameroon Dolphin, the Gulf of Guinea Population of Sousa teuszii (Kükenthal, 1892). ISRN Biodiversity 2014: 819827. http://dx.doi.org/10.1155/2014/819827
Gulf of Mannar (between India and Sri Lanka)
Avelin, Mary, Mary, Vitalina, Lorella, Araceli and Matias, Jonathan R. (2009). Rediscovery of naturally occurring seagrape Caulerpa lentillifera from the Gulf of Mannar and its mariculture. Current Science 97(10): 1418-1420.
Haiti
Skog, L. E. and Talpey, T. E. (1971). Rediscovery of Gesneria humilis. Gloxinian 21(3): 7-9.
Honduras
Ordoñez-Mazier DI, Ávila-Palma HD, Soler-Orellana JA, Ordoñez-Trejo EJ, Martínez M, Turcios-Casco MA. (2020). Rediscovery of Vampyressa thyone (Chiroptera, Phyllostomidae) in Honduras: updated distribution and notes on its conservation. Neotropical Biology and Conservation 15(3): 379-390. https://doi.org/10.3897/neotropical.15.e53766
Hungary
Erzberger, Peter et al. (2015). Revision of the Red List status of Hungarian bryophytes 1. new occurrences of species previously thought to be regionally extinct or without recent data. Studia Botanica Hungarica 46(2): 15-53. [Ulota hutchinsiae, Brachythecium geheebii, Fontinalis hypnoides, Isopterygiopsis pulchella, Riccardia chamedryfolia]
Kontschán, Jenő, Attila, Takács and Kőszegi, Klaudia. (2024). Rediscovery of Livilla radiata (Foerster, 1848) after 130 years in Hungary, with notes to the Fabaceae-associated jumping plant lice in Hungary (Insecta: Psylloidea). Ecologica Montenegrina 71: 139-143.
Kontschán, Jenő, Kiss, E. and Ripka, G. (2019). Rediscovery of three rare spider mites in Hungary (Acari: Tetranychidae). Növényvédelem 55(6): 249-252. [Abstract]
Kontschán, Jenő, Kiss, E. and Ripka, G. (2020). Rediscovery of Cenopalpus lineola (Canestrini and Fanzago, 1876) in Hungary (Acari Tenuipalpidae). Acta Phytopathologica et Entomologica Hungarica 55(1): 103-114.
Koszka, Attila and Papp, Viktor. (2020). Aurantiporus croceus, a flagship species of the European fungal conservation is re-discovered after half century in Hungary. Acta Biologica Plantarum Agriensis 8(1): 40-52.
Kovács, T. & A. Ambrus, A. (2001). Ephemeroptera, Odonata and Plecoptera larvae from the rivers of Rába and Lapincs (Hungary).- Folia Historico-Naturalia Musei Matraensis 25: 145-162. [Palingenia longicauda]
Lengyel, A. (2009). New occurrence of Botrychium matricariifolium (Retz.) A. Braun ex W.D.J. Koch in Hungary. Acta Botanica Hungarica 51(1-2): 99-104.
Schmotzer, András and Táborská, Jana. (2016). Rediscovery of Campanula macrostachya Waldst. & Kit. Ex Willd. in the Bükk Foothills. Acta Biologica Plantarum Agriensis 4: 72-88.
Seres, G., Romsauer, J. and Merkl, O. (2014). Rediscovery of Parazuphium chevrolatii praepannonicum in Hungary (Coleoptera: Carabidae). Folia Entomologica Hungarica 75: 15-19.
India
A. J., Robi and Udayan, P. S. (2020). A re-investigation on the identity of Litsea nigrescens (Lauraceae) in the Western Ghats of India. Rheedea 30(2): 270-277.
A. R., Viji and Pandurangan, A. G. (2015). On the occurrence of little known sedge, Carex walkeri from Nilgiri Mountains, Western Ghats, India. Int. J. Pure App. Biosci. 3(2): 105-107.
Aishwarya, K. et al. (2015). A new species of Boesenbergia and rediscovery of B. rotunda (Zingiberaceae) from India. Phytotaxa 197(3): 186-196.
Akhilesh, K. V. et al. (2011). Rediscovery and description of the quagga shark, Halaelurus quagga (Alcock, 1899) (Chondrichthyes: Scyliorhinidae) from the southwest coast of India. Zootaxa 2781: 40-48. [Abstract]
Alam, A., S. Vats & K.K. Behera (2012). Exormotheca ceylonensis Meijer-A Threatened Liverwort in India, rediscovered in Palni Hills, Tamil Nadu. Journal of Threatened Taxa 4(5): 2593–2595.
Arockianathan, Samson et al. (2014). Calliophis bibroni (Bibroni Coral Snake): Rediscovery in Mudumalai Tiger Reserve, South India. Herpetological Bulletin 127: 35-36.
Arumugam, S., K.S. Kumar, B. Karthik & V. Ravichandran (2019). Cordia difusa K.C. Jacob, the Kovai Manjack (Boraginaceae): a highly threatened stenoendemic species from Coimbatore City, Tamil Nadu, India.Journal of Threatened Taxa 11(14): 14870-14875.
Asthana, G. and Saxena, M. (2011). Lejeunea kodamae Ikegami & Inoue new to India, with a note on the rediscovery of L. bidentula Herzog. Journal of Bryology 33(1): 89-92. https://doi.org/10.1179/1743282010Y.0000000005
Augustine KT. 2003 Rediscovery of Phyllanthus rotundifolius Klein ex Willd., Euphorbiaceae, after 101 years (1899-2000) in Kanchipuram District, Tamil Nadu, South India. J. Bombay Nat. Hist. Soc. 99. (3): 562-564.
Baidya, Sarika et al. (2023). Rediscovery of Lethe tristigmata Elwes, 1887 (Lepidoptera: Nymphalidae: Satyrinae). ZooNova 26: 1-9.
Balan, A. P. and Predeep, S. V. (2016). Note on the occurrence of Dunbaria punctata (Leguminosae: Papilionoideae), a little-known legume from the Western Ghats, India. Rheedea 26(2): 157-160.
Bhatt, Harish, Kumar, Shankar and Singh, Paramjit. (2020). Rediscovery of small silverfork, Lethe jalaurida (Insecta: Lepidoptera: Nymphalidae) from Garhwal, Uttarakhand, India. Bionotes 22(4): 198-200.
Borah, Dipankar et al. (2020). Rediscovery of Tupistra stoliczkana (Asparagaceae) in northeastern India and the identity of T. ashihoi. Phytotaxa 443(2): 207-210. https://doi.org/10.11646/phytotaxa.443.2.8 [Abstract] [specific name is actually stoliczana]
Borah, M. M. and Bordoloi, S. (2001). Rediscovery of Bufo macrotis Boulenger (Amphibia: Anura) in Arunachal Pradesh, India. Zoos' Print Journal 16(8): 574.
Chakrovorty, Arnob et al. (2023). A note on the rediscovery of the sand wasp Bembecinus proximus (Handlirsch) from an urban habitat. Indian Journal of Entomology Online [2023]: e23148. doi:10.55446/IJE.2023.1148
Choudhury, A., Chandiramani, S. S. and Naik, R. N. (1999). White-winged wood duck rediscovered in Namdapha. Tigerpaper 26(2): 27-28. [Asarcornis scutulata]
Choudhury, Kushal. (2010). Rediscovery of two rare butterflies Papilio elephenor Doubleday, 1845 and Shijimia moorei Leech, 1889 from proposed Ripu-Chirang Wildlife Sanctuary, Assam, India. Journal of Threatened Taxa 2(4): 831-834. [local rediscovery of Shijimia moorei, global rediscovery of Papilio elephenor]
Dahal, S., Sharma, T. P. and Borthakur, S. K. (2017). Rediscovery of Aconitum novoluridum (Ranunculaceae) from Sikkim Himalaya, India. Rheedea 27(2): 116-118. https://dx.doi.org/10.22244/rheedea.2017.27.2.23
Dewan. S., B.K. Acharya & S. Ghatani (2018). A new record of the lesser-known buterfy Small Woodbrown Lethe nicetella de Nicéville, 1887 (Lepidoptera: Nymphalidae: Satyrinae) from Khangchendzonga Natonal Park, Sikkim, India. Journal of Threatened Taxa 10(6): 11775–11779.
Dey, D., M. Baruah, N. Devi & J.N. Borah (2020). Re-collection of the Luminous Lantern Flower Ceropegia lucida Wall. (Apocynaceae) from Assam, India. Journal of Threatened Taxa 12(1): 15212–15215.
Dey, Sentu Kumar and Maity, Debabrata. (2015). Rediscovery of Pseudoyoungia simulatrix (Cichorieae-Asteraceae) from India and its taxonomic history. Rheedea 25(1): 31-35.
Dey, Sentu Kumar and Maity, Debabrata. (2016). The rediscovery of Youngia atripappa (Babc.) N. Kilian in India after a century; its circumscription and nomenclature and the lectotypification of Crepis atripappa Babc. Taiwania 61(4): 362-368.
Dey, D., Yadav, S. R. and Devi, N. (2021). Rediscovery of Aponogeton lakhonensis A. Camus (Aponogetonaceae): a long-lost aquatic plant of India. Journal of Threatened Taxa 13(11): 19632-19635. https://doi.org/10.11609/jott.7464.13.11.19632-19635
Gautum, Ajay Kumar and Avasthi, Shubhi. (2017). Discovery of Puccinia tiliaefolia (Puciniales) in northwestern Himalayas, India. Polish Botanical Journal 62(1): xx-xx.
Ghosh, Chandra, Paul, T. K. and Das, Abhaya Prasad. (2013). Rediscovery of Hibiscus fragrans Roxburgh (Malvaceae) from Jaldapara National Park in Duars of West Bengal, India. Pleione 7(2): 531-537.
Jalal, J. S. and Pangtey, Y. P. S. (2011). Rediscovery of Vanda alpina, a rare epiphytic orchid in Western Himalaya, India. Richardiana 9(4): 173-178.
James, D. B. (1966). Studies on Indian Echinoderms-I Rediscovery of the Echinoid, Breynia verdenburgi Anderson from Andaman Sea, with an emended description. Journal of Marine Biological Association of India 8: 76-81.
Joe, A., Sreejith, P. E., Ashfak, O. A. and Sabu, M. (2014). Regarding the identity, rediscovery and taxonomic history of Musa nagensium (Musaceae) from India. Rheedea 24(1): 5-11. https://dx.doi.org/10.22244/rheedea.2014.24.01.02
Kavade, S., S. Deokule, P. Lakshminarasimhan, P. Diwakar & S. Punekar (2012). Belosynapsis vivipara (Dalzell) C.E.C. Fisch. (Commelinaceae), a vulnerable spiderwort, rediscovered after sixteen decades from Maharashtra, India. Journal of Threatened Taxa 4(6): 2660-2663.
Kishor, Rajkumar. (2012). Rediscovery of Kaempferia marginata (Zingiberaceae) after a lapse of 100 years from India. Rheedea 22(1): 28-31. https://dx.doi.org/10.22244/rheedea.2012.22.01.08
Kumar, Mohd. Umar, Mochi, Shakir Ahmad and Riyaz, Muzafar. (2022). Rediscovery, distribution and taxonomic description of Astragalus trichocarpus Benth. (Fabaceae) from the Northwestern Himalayas - Kashmir, India. Species 23(71): 169-172.
Kumar, Rahul and Sharma, Ajay Kumar. (2022). Rediscovery of Campanacella hamiltonella (Westwood, 1867) (Neuroptera: Mantispidae) from India after 155 years. Uttar Pradesh Journal of Zoology 43(5): 78-85. [Abstract]
Kumar S, Singh RS, Singh P, Kumar S. (2019). Rediscovery of butterflies Arhopala bazalus Hewitson, 1862 and Catapaecilma major Druce, 1895 from Uttarakhand, India. Journal of Entomology and Zoology Studies 7(1): 125-128.
Kumar, Shankar et al. (2020). Rediscovery of Tiger hopper Ochus subvittatus (Insecta: Lepidoptera: Hesperiidae) from Uttarakhand, India. Bionotes 22(4): 216-217.
Kumar, Y. (1992). 'Extinct' orchid rediscovered. Current Science 62(8): 547-548. [Bulbophyllum rothschildianum]
Kunte, K. and Manthey, U. (2009). Rediscovery of Japalura sagittifera (Sauria: Agamidae)from the Eastern Himalayas, Arunachal Pradesh: An addition to the Indian Herpetofauna. Sauria 39(2): 42-55.
S. N. Litvinchuk, D. A. Melnikov, L. J. Borkin, S. Hofmann. (2019). Rediscovery of the High Altitude Lazy Toad, Scutiger occidentalis Dubois, 1978, in India. Russian Journal of Herpetology 26(1): 17-22. [Abstract]
Maity, Rohan et al. (2022). Rediscovery of Phlogacanthus jenkinsii (Acanthaceae) from India after 140 years. Phytotaxa 573(2): 286-292. https://doi.org/10.11646/phytotaxa.573.2.8
Majumder, Tamoghno, Kairy, Susmita, Shekhar, Shashank, E., Sankarganesh and Roy, Kusal. (2026). Rediscovery and ecological notes on Amida vietnamica Hoang, 1982 (Coleoptera: Coccinellidae): a little-known ladybird beetle from India. International Journal of Tropical Insect Science. https://doi.org/10.1007/s42690-026-01746-1
Mao, A. A., Krishna, Gopal and Jalal, Jeewan Singh. (2022). Notes on recollection of Primula sherriffiae W.W.Sm. after a gap of 74 years from India with comments on the typification of the name. Phytotaxa 571(2): 234-238. https://doi.org/10.11646/phytotaxa.571.2.10
Mathew, Jose and George, K. V. (2013). Rediscovery of Arisaema jacquemontii Blume (Araceae) from Achankovil Valley of Southern Western Ghats, India. NeBIO 4(6): 96-97.
Meena, S.S., A. Tripathi, V.K. Koli & M.A. Awan. (2021). Rediscovery of the rare Desert Grizzled Skipper Spialia doris evanida Butler, 1880 (Hesperiidae: Pyrginae) from the Thar Desert, Rajasthan, India. Journal of Threatened Taxa 13(3): 18042-18044. https://doi.org/10.11609/jott.6348.13.3.18042-18044
Mir, Aabid Hussain, Upadhaya, Krishna, Odyuo, Nripemo and Tiwari, B. K. (2017). Rediscovery of Magnolia rabaniana (Magnoliaceae): a threatened tree species of Meghalaya, northeast India. Journal of Asia-Pacific Biodiversity 10(1): 127-131. http://dx.doi.org/10.1016/j.japb.2016.10.004
Moaakum and Dey, S. (2013). Rediscovery of Hedychium longipedunculatum A.R.K. Sastry & D.M. Verma [Zingiberaceae] from Nagaland, India. Pleione 7(1): 290-294.
Mohanty, Biswaprajna et al. (2019). First record of a Leucosid crab Paranursia abbreviata Bell, 1855 from Devi estuary, Odisha Coast, India. Indian Journal of Geo Marine Sciences 48(1): 117-119.
Mondal, Sujit et al. (2018). Areca triandra Roxb. ex Buch.-Ham. (Arecaceae): new record for West Bengal, India. Plant Archives 18(2): 1700-1702.
Muansanga, Lal et al. (2021). Rediscovery of Doria’s Foam-nesting Treefrog, Chirixalus doriae Boulenger 1893 (Anura: Rhacophoridae), from India. Reptiles & Amphibians 28(1): 79-81.
Nair, N. C. and Srinivasan, S. R. (1983 "1982"). On the rediscovery of Koilodepas calycinum Bedd. (Euphorbiaceae) and Holcolemma canaliculatum (Nees ex Steud.) Stapf. ex Hubbard (Poaceae) from south India. Bull. Bot. Surv. India 24(1-4): 241-242. [Holcolemma canaliculatum]
Naithani, H. B. (2021). Rediscovery of Strobilanthes pterygorrhachis C.B. Clarke (Acanthaceae) from India. The Indian Forester 147(12): 1233. http://dx.doi.org/10.36808/if%2F2021%2Fv147i12%2F166973
Nandikar M D, Gurav R V and Umesh D. (2011). Rediscovery of Murdannia striatipetala (Commelinaceae), a little known species from Southern India with a note on its identity and distribution. Journal of Bombay Natural History Society 108(1): 67-68.
Panday, Samiran and Sinha, B. K. (2012). Rediscovery of Campanumoea javanica Blume- a rare climber of Campanulaceae from Phawngpui National Park in Mizoram, India. Pleione 6(1): 241-243.
Ragavan, P. et al. (2016). The rediscovery of Brownlowia tersa (L.) Kosterm. (Malvaceae), from the Andaman Islands, India — a Near Threatened mangrove species. Check List 12(3): 1897.
Rai, S., K.D. Bhutia & K. Kunte (2012). Recent sightings of two very rare butterflies, Lethe margaritae Elwes, 1882 and Neptis nycteus de Nicéville, 1890, from Sikkim, eastern Himalaya, India. Journal of Threatened Taxa 4(14): 3319–3326. [local rediscovery of both taxa]
Rajkumar, Muthusamy, Saravanan, Raju and Maheswarudu, Gidda. (2018). Rediscovery of rare porcellanid crab Pseudoporcellanella manoliensis Sankarankutty, 1961, from Palk Bay, Tamil Nadu, India. Indian Journal of Geo Marine Sciences 47(11): 2182-2185.
Rawat, N. and Acharya, B. K. (2024). Rediscovery of rare Anisoptera, Camacinia harterti, Karsch, 1890 (Odonata: Libellulidae) from Sikkim Himalaya, India. Int J Trop Insect Sci 44(2): 989-993. https://doi.org/10.1007/s42690-024-01182-z
Reddy, C.S., S.M.M. Javed, F. Tampal, C. Pattanaik & V.S. Raju (2010). Rediscovery of Chamaesyce sebastinei (Binojk. & N.P. Balakr.) V.S. Raju (Angiosperms: Euphorbiaceae) a little-known rare species from Andhra Pradesh, India. Journal of Threatened Taxa 2(9): 1160-1161.
Sadasivan, Kalesh et al. (2021). Rediscovery of Martin’s Duskhawker Anaciaeschna martini (Selys, 1897) (Odonata: Aeshnidae) from Western Ghats, peninsular India, with notes on its current distribution and oviposition behavior. Journal of Threatened Taxa 13(1): 17543-17547. https://doi.org/10.11609/jott.6301.13.1.17543-17547
Saji Kumar, K K and Rajeeshkumar, M P and Laxmilatha, P and Manjebrayakath, Hashim and Venkatesan, V and Saravanane, N. (2022). Rediscovery of Ram’s horn squid, Spirula spirula (Cephalopoda: Spirulidae), from the Arabian Sea. Journal of the Marine Biological Association of India 64(1): 69-73.
Sarania, B., Devi, A., Kumar, A., Wang, K. and Rakshit, K. (2015). A record of Scutiger nyingchiensis Fei, 1977 (Amphibia: Anuran: Megophryidae) in the Eastern Himalaya, North East India. Current Science 109(3): 413-414.
Sarode, B.V., N.U. Joshi, P.P. Pansare & H.V. Ghate (2018). A record afer 52 years, and additonal descripton of the emesine assassin bug Emesopsis nubila (Hemiptera: Reduviidae: Emesinae) from western India. Journal of Threatened Taxa 10(9): 12282-12285.
Sharma, A., N. Singh & P. Kumar (2019). Rediscovery of Calanthe davidii (Orchidaceae) after 11 decades in the western Himalaya, India. Journal of Threatened Taxa 11(10): 14368-14372.
Singhamahapatra, Aniruddha and Roy, Subhajit. (2023). First record of the rare moth Amana angulifera Walker, 1855 (Lepidoptera: Epicopeiidae) from India since the 19 th century. Cuadernos de Biodiversidad 64: 19-23. https://doi.org/10.14198/cdbio.22415
Singh, Arun P. (2013). Rediscovery of the Empress, Sasakia funebris Leech (Lepidoptera: Nymphalidae: Nymphalinae: Apaturini) after 88 years in India. Journal of Threatened Taxa 5(10): 4514-4516.
Singh, Bikarma and Bedi, Yashbir Singh. (2016). Rediscovery, Taxonomic History and Extended Enumeration of Haematocarpus validus Bakh.f. ex Forman (Menispermaceae) to Indo-Myanmar Biodiversity Hotspot. National Academy Science Letters. DOI: 10.1007/s40009-016-0483-8 [Abstract]
Singh, D. K. and Majumdar, Shuvadeep. (2017). Notes on Scarcely Collected Indian Liverworts III. Plagiochila kurzii (Plagiochilaceae, Marchantiophyta). Indian Journal of Forestry 40(2): 155-158.
SINGH, W., HANDIQUE, P. & DEVI, H. (2016). Musa sikkimensis Kurz (Musaceae): A note on the rediscovery from Manipur, India. Folia Malaysiana, 17, 47-53.
Sreebha, R., Kariyappa, K. C. and Daniels, A. E. D. (2016). Rediscovery of a long-lost moss Fissidens serratus var. serratus in the Western Ghats of India. Acta Botanica Hungarica 58(1-2): 183-186. [Abstract]
Srinivasulu, C., D. Venkateshwarlu & M. Seetharamaraju (2009). Rediscovery of the Banded Krait Bungarus fasciatus (Schneider 1801) (Serpentes: Elapidae) from Warangal District, Andhra Pradesh, India. Journal of Threatened Taxa 1(6): 353-354.
Swamy, J. and Arumugam, S. (2021). Sporobolus tetragonus (Poaceae: Chloridoideae), an addition to the flora of South India. Rheedea 31(4): 296-299. https://dx.doi.org/10.22244/rheedea.2021.31.04.06
Venugopal, N. and Pamidimarri, Stella. (2015). Rediscovery of critically endangered Gymnocladus assamicus Kanjilal ex P.C. Kanjilal (Leguminosae: Caesalpinioideae) from West Khasi Hills District of Meghalaya. Pleione 9(1): 186-192.
Psydrax salicifolia [global rediscovery?]
Sarcandra chloranthoides (global rediscovery?): http://www.vattakanalconservationtrust.org/Home/activities
Brettus cingulatus: https://www.nyoooz.com/news/mangalore/819585/jumping-spider-springs-back-after-122-years/
Lumnitzera racemosa: https://www.hindustantimes.com/cities/very-rare-species-of-mangroves-rediscovered-after-63-years-report/story-D1ujM1QMVZ7RGaNhdFAO0O.html
Andaman Islands
Gupta, Stutee et al. (2002). Rediscovery of Podocarpus wallichianus: A rare gymnosperm from tropical rain forests of Great Nicobar Island. Current Science 87(3): 806-807.
Karthigeyan, K., Sumathi, R. and Jayanthi, J. (2012). Recollection of a rare epiphytic orchid Taeniophyllum filiforme J.J. Sm. (Orchidaceae) after a lapse of 135 years from South Andaman Islands, India. Journal of Threatened Taxa 4(15): 3433-3435. https://doi.org/10.11609/JoTT.o2851.3433-5
Mathew, Sam P. (2013). Thunia alba (Lindl.) Reichb.F. a rare wild ornamental orchid from the Andaman Islands in the Bay of Bengal. Orchid Digest 77(3): 150-151.
Ramana, M. V., Venu, P. and Sanjappa, M. (2012). The rediscovery of Polyalthia crassa (Annonaceae) from Andaman and Nicobar Islands, India. Rheedea 22(1): 5-8. https://dx.doi.org/10.22244/rheedea.2012.22.01.02
Andhra Pradesh
Ramana, S. P. Venkata and Harinth, P. (2022). Rediscovery of the endangered spider tarantula Poecilotheria formosa from Sri Lankamalleswara reserve forest - Southern Andhra Pradesh - India. Journal of Applied Entomologist 2(3): 33-36.
Arunchal Pradesh
KHANDAL D., MORE S., KATARIA G. & S.S. KAMBALE. (2018). Ceropegia lucida–rediscovery and new distribution record for Arunachal Pradesh, India. Current Science 113(11): 2077.
Assam
Baruah, S. et al. (2013). Notes on two interesting angiosperms from Assam, India. Pleione 7(2): 401-405.
Gogoi, Khyanjeet. (2023). Rediscovery of Calanthe nana (Orchidaceae) and new addition to the flora of Assam. Richardiana 7: 132-139.
Kemprai, Kapil Kumar and Borah, Souravjyoti. (2026). Rediscovery of Strobilanthes panichanga and discovery of Strobilanthes parryorum (Acanthaceae): two endangered species from the hills of Dima Hasao, Assam, India. Nordic Journal of Botany 2026: e05101. https://doi.org/10.1002/njb.05101
Pagag, K. and Borthakur, S. K. (2012). Rediscovery of Blyxa japonica (Miquel) Maximowicz ex Ascherson et Gürke - a hydrophyte from Lakhimpur district of Assam, India. Pleione 6(2): 435-437.
Gujarat
Patel, Mitesh and Reddy, Mandadi Narsimha. (2018). Equisetum ramosissimum Desf. Subsp. debile (Roxb. ex Vaucher) Hauke – is really extinct from Gujarat, India? International Journal of Botany Studies 3(4): 21-24.
Rana, Karan G. et al. (2015). Rediscovery of Olax nana (Olacaceae) for Gujarat state. J. Econ. Taxon. Bot. 39(2): 297-299.
Jammu and Kashmir
Adhikari, BS, A. Kunzes, JS Jalal and GS Rawat. (2013). Rediscovery of Dactylorhiza kafiriana, a lesser known orchid from Suru valley, Jammu and Kashmir, India. Richardiana 8: 220-225.
Dar, Afaq Ahmad et al. (2022). Detailed Taxonomic Redescription, Distribution, and Rediscovery of the Birch Emperor Moth, Rinaca lindia (Moore 1865) (Bombycoidea: Saturniidae) from Jammu & Kashmir, India, After a 38-Year Hiatus. The J. of the Lepidopterists' Society 76(3): 203-206. https://doi.org/10.18473/lepi.76i3.a5
Sharma, Shakha and Sharma, Neeraj. (2020). On the rediscovery and new additions of Skippers (Lepidoptera: Papilionoidea: Hesperiidae) from Jammu and Kashmir, India. Revista Chilena de Entomología 46(4): 591-599. [Pseudocoladenia fatih]
Sheikh, Taslima et al. (2022). Rediscovery of white-bordered copper, Lycaena panava (Westwood, 1852) (Lepidoptera: Lycaenidae) from Union Territory of Jammu and Kashmir, India. Life Sciences Leaflets 144: 1-5.
Sheikh, T. and Mishra, S. (2023). A note on the reconfirmation of tawny mime, Papilio agestor Gray, 1831 (Lepidoptera: Papilionidae) from union territory of Jammu and Kashmir, India after 115 years. Munis Entomology & Zoology 18(1): 629-633
Singh, Sumit and Sheikh, Taslima. (2021). Rediscovery of Popinjay, Stibochiona nicea (Gray, 1846) (Lepidoptera: Nymphalidae: Nymphalinae) from Union Territory of Jammu and Kashmir, India. Revista Chilena de Entomología 47(3): 497-499. http://dx.doi.org/10.35249/rche.47.3.21.07
Karnataka
Sant, N., Shelke, V. and Shelke, S. (2016). Rediscovery of the Broad-tailed Grassbird Schoenicola platyurus at Belgaum, Karnataka. Indian BIRDS 12 (4&5): 145-147.
Kashmir
Riyaz, Muzafar and Sivasankaran, K. (2022). REDISCOVERY AND DISTRIBUTION OF ORETA VATAMA LUCULENTA WATSON, 1967 (LEPIDOPTERA: DREPANIDAE) FROM THE PIR PANJAL VALLEY, NORTH-WESTERN HIMALAYAS, KASHMIR, INDIA. Mun. Ent. Zool. 17(1): 619-621.
Madhya Pradesh, India
Lohit, Tushar A. et al. (2022). Nomenclatural notes and rediscovery of Viola cinerea var. stocksii (Violaceae) from Chambal ravines of Madhya Pradesh, India. Phytotaxa 566(2): 209-218. https://doi.org/10.11646/phytotaxa.566.2.5
Maharashtra
Sathaye, G., Ranade, S. and Ghate, H. V. (2022). Rediscovery of an interestng preying mants Deiphobella latceps (Mantodea: Rivetnidae) from Maharashtra, India. Journal of Threatened Taxa 14(11): 22190-22194. htps://doi.org/10.11609/jot.8049.14.11.22190-22194
Manipur
Dhatchanamoorthy, Narayanasamy, Ravikumar, Kaliamoorthy and Balachandran, Natesan. (2023). Rediscovery of Polygonatum tessellatum F. T. Wang & Tang in Manipur, north-eastern India. Biodiv. Res. Conserv. 70. [Abstract]
NGANGBAM, R. D., DEVI, N. P., DEVI, M. H. & SINGH, P. K. 2019. Rediscovery of Aldrovanda vesiculosa L. (Droseraceae), an endangered plant, from Manipur in India after six decades, with studies on micromorphology and physico-chemistry of water. Reinwardtia 18(2): 71-80.
Soibam, Baleshwor Singh, Huidrom, Harmenn and Irungbam, Jatishwor Singh. (2016). A century later: Tricolored Pied Flat Coladenia indrani uposathra Fruhstorfer, 1911 (Hesperiidae: Pyrginae) and Crenulate Oakblue Apporasa atkinsoni Hewitson, 1869 (Lycaenidae: Theclinae) reported from Manipur, India. Journal of Threatened Taxa 8(7): 9030-9033.
Meghalaya
Mahesh, Y., Lytan, R. & Kottaimuthu, R. (2026). Rediscovery of Chamaegastrodia vaginata (Hook.f.) Seidenf. (Orchidaceae: Goodyerinae) after a gap of 175 years from its type locality. Vegetos. https://doi.org/10.1007/s42535-025-01592-5
Shankar, U. (2021). Lectotypification and recollection of Bulbophyllum crabro in Meghalaya after 125 years. Rheedea 31(3): 211-217. https://dx.doi.org/10.22244/rheedea.2021.31.03.13
Sharma, Sachin et al. (2021). Rediscovery of Hedge Cupid, Bothrinia chennellii (de Nicéville, 1884) (Lepidoptera: Lycaenidae) to Meghalaya, Northeastern India. Revista Chilena de Entomología 47(3): 493-496.
Odisha
Mahata, A., Jena, S. K. and Palita, S. K. (2019). First record in 129 years of the Tamil Treebrown Lethe drypets todara Moore, 1881 (Lepidoptera: Nymphalidae: Satyrinae) from Odisha, India by fruit-baitng. Journal of Threatened Taxa 11(15): 15047-15052.
Tripathy, A. (2020). Rediscovery of Cyclopodia sykesii, an ectoparasite on the Indian Flying Fox from Udayagiri Range, Odisha, India. Vet Brief #12, In: Zoo’s Print 35(6): 36-38.
Rajasthan
Dookia, S., G. Singh & R. Mishra (2017). Re-sighting record of Fulvous Leaf-nosed Bat Hipposideros fulvus Gray, 1838 (Mammalia: Chiroptera: Hipposideridae) from Thar Desert, Rajasthan, India. Journal of Threatened Taxa 9(1): 9764-9767.
Sikkim
Nautiyal, D. C., Sharma, S. K. and Pandit, M. K. (2009). Notes on the taxonomic history, rediscovery and conservation status of two endangered species of Ceropegia (Asclepiadaceae) from Sikkim, India. Journal of the Botanical Research Institute of Texas 3(2): 815-822. [Ceropegia lucida]
Pradhan, D. K. (2021). Recollection of Galeola falconeri Hook. f. (Orchidaceae) from Sikkim Himalaya after 123 years. Pleione 15(2): 246-248.
Tamil Nadu
Britto, S. J. (2002). Rediscovery of Crotalaria orixensis Rottler ex Willd. (Papilionoideae) after 90 years - a little known and rare species in Tamil Nadu. J. Econ. Taxon. Bot. 26(3): 583-585.
Ganesh, S. R. and S. R. Chandramouli. (2010). Rediscovery of Hemidactylus scabriceps (Annandale, 1906) (Reptilia: Sauria: Gekkonidae) from Eastern Tamil Nadu, India. Russ. J. Herpetol. 17(1): 70-74.
MosaChristas, Kithiyon, Karthick, Ravikumar, Kowsalya, Elumalai and Jaquline, Chinna Rani I. (2021). Musa kattuvazhana (Musaceae): Rediscovery and additional notes on a critically endangered species from Western Ghats of Tamil Nadu, India. Feddes Repertorium 132(3): 263-268. https://doi.org/10.1002/fedr.202000034
Ravikumar, Kaliamoorthy, Sankar, R. Vijaya and Prabhakaran, V. (2001). Utleria salicifolia Bedd. ex Hook.f. (Periplocaceae) A Monotypic, Endemic and Critically Endangered, Red Listed Medicinal Plant relocated from its type locality. MyForest 37(2): 495-498.
Tejavathi, D. H., Sumalatha, B. S., Nijagunaiah, R., Anitha, P. and Gayatramma, K. (2019). Studies on the Factors that Restrict Sexual Reproduction and Fruit Setting in Memecylon flavescens: An Endangered Taxon. Journal of Tropical Forestry and Environment 9(2): 101-107.
Uttar Pradesh
Kumari, Priyanka and Sheikh, Taslima. (2021). A note on the rediscovery of the Redspot butterfly Zesius chrysomallus Hübner, 1819 (Lepidoptera: Lycaenidae: Theclinae) from Uttar Pradesh State, with a new larval host plant record for India. Revista Chilena de Entomología 47(2): 399-404.
Uttarakhand
Kasana, Shruti, Uniyal, P. L. and Pandey, Arun K. (2019). Rediscovery of Saussurea andryaloides (Asteraceae: Cardueae) from Uttarakhand, India. Rheedea 29(1): 116-118.
Kumar, Shankar et al. (2019a). Rediscoveries of dusky yellow-breasted flat Gerosis phisara Moore (Lepidoptera: Hesperiidae) and angled Pierrot Caleta decidia hewitson (Lepidoptera: Lycaenidae) from Uttarakhand. Journal of Entomology and Zoology Studies 7(1): 45-48.
Kumar, Shankar et al. (2019b). Rediscovery of butterflies Arhopala bazalus Hewitson, 1862 and Catapaecilma major Druce, 1895 from Uttarakhand, India. Journal of Entomology and Zoology Studies 7(2): 864-867.
Kumar, Shankar et al. (2020). Rediscovery of white-striped snow flat Tagiades cohaerens Mabille, 1914 (Lepidoptera: Hesperiidae) from Uttarakhand, India. Journal of Entomology and Zoology Studies 8(1): 1431-1433. [Tagiades cohaerens cynthia]
West Bengal
Apurba Chakraborty, Sayan Tripathi & Bidyut B. Bhattacharya. (In Press, 2018). Rediscovery of the Spoon-billed Sandpiper Calidris pygmaea on the coast of West Bengal, India. Indian BIRDS 14: 83-84.
Chowdhury, S. and Soren, R. (2009). Rediscovery of Small Salmon Arab Colotis amata Fabricius (Lepidoptera: Pieridae) from saline and semi-saline areas of West Bengal, India. Journal of Threatened Taxa 1(6): 351-352.
Deuti, K., Biswas, S., Ahmed, M. F. and Dutta, S. K. (2000). Rediscovery of Chirixalus simus Annandale, 1915 (Anura: Rhacophoridae) from Assam and West Bengal, eastern India. Hamadryad 25(2): 215-217.
Jash, Santayan and De, Asit Baran. (2024). On the Maravalia echinulata (Niessl ex Rabenh.) Ono (Pucciniales: Chaconiaceae) with reference to its host range and distributon. Journal of Threatened Taxa 16(12): 26283-26290. https://doi.org/10.11609/jot.6876.16.12.26283-26290
Indonesia
Auliya, M. (2002). Rediscovery of the Indochinese Rat Snake Ptyas korros (Schlegel, 1837) (Serpenthes: Colubridae) in Borneo. The Raffles Bulletin of Zoology 50(1): 197-198.
Putra, Ivan Permana, Hermawan, Rudy and Maulana, Indra. (2022). Re-discovery of the rare poisonous mushroom Podostroma cornu-damae in Indonesia. Biharean Biologist 16(2): 60-64.
Putra, Ivan Permana and Nurhayat, Oktan Dwi. (2024). The Recent Collection of Climacodon roseomaculatus (Hydnum roseomaculatum Henn. & E. Nyman) from The Type Locality (Indonesia). Biology, Medicine, & Natural Product Chemistry 13(2): 585-589.
Trainor, Colin R. (2002). An expedition to Damar Island, South West Maluku, Indonesia. Bulletin of the Oriental Bird Club 36: 18-23. [Chrysococcyx rufomerus]
Soejono. (2014). Rediscovery of a remnant habitat of the critically endangered species, Hopea sangal, in Pasuruan District, East Java, Indonesia. Biodiversitas 15(2): 236-239.
Sulawesi
Damayanto, I Putu Gede P. and Riastiwi, Indira. (2019). Balanophora papuana Schltr. (Balanophoraceae), a Neglected Holoparasite Species: Rediscovery for Indonesia. Proceedings of the 3rd SATREPS Conference, Bogor, 22 November, 2018, pp. 102-110.
Iran
Bagheri, Ali et al. (2018). Rediscovery of Astragalus saganlugensis (Fabaceae, Galegeae) in Iran after 184 years. Phytotaxa 350(3): 297-299. [Abstract]
Gholipour, Abbas. (2013). Rediscovery of Acorus calamus (Acoraceae) in Iran. Journal? [Abstract]
Jalilian, N., Nemati Paykani, M. and Soleimani, F. (2021). Rediscovery of Dionysia bornmuelleri (Pax) Clay (Primulaceae) in West Iran. Iranian Journal of Botany 27(2): 98-102. doi:10.22092/ijb.2021.355470.1333
Kidov, A. A. (2021). Rediscovery of the Persian dwarf snake (Eirenis persicus, Ophidia, Colubridae) in Qazvin Province, Iran. Zoologicheskiy Zhurnal 100(6): 690-692. [Abstract]
Najafi-Majd, Elnaz and Kaya, Uğur. (2013). Rediscovery of the Lake Urmia newt, Neurergus crocatus Cope, 1862 (Caudata: Salamandridae) in northwestern Iran after 150 years. Amphibian and Reptile Conservation 6(4): 36-41.
Schneider, T., Schneider, E., Schneider, J. and Müller, O. (2014). Rediscovery of Cordulegaster vanbrinkae in Iran (Odonata: Cordulegastridae). Odonatologica 43: 25-34.
Shafiei, Soheila et al. (2014). Rediscovery of Maynard's Longnose Sand Snake, Lytorhynchus maynardi, with the geographic distribution of the genus Lytorhynchus Peters, 1863 in Iran. Zoology in the Middle East. DOI:10.1080/09397140.2014.994309 [Abstract]
Ireland
Alexander, K. N. (2009). The rediscovery of some of Ireland's rarest wood-decay beetles (Coleoptera: Lymexylidae, Phloiophilidae, Ciidae, Tetratomidae, Melandryidae). The Irish Naturalists’ Journal 30(2): 130-133.
Bannister, P. (1996). A rediscovered site for Erica erigena R. Ross (Ericaceae) on Achill Island, County Mayo (v.c. H27), Ireland. Watsonia 21(1): 99-101.
O'Reilly, Noreen and Boparai, Jaswinder S. (2024). Rediscovery of Banchus crefeldensis Ulbricht (Hymenoptera: Ichneumonidae: Banchinae) in Ireland. Entomologist's Monthly Magazine 160(4): 287-292. https://doi.org/10.31184/M00138908.1604.4249
Israel
Vaginsky, E. (1990). The Bat Miniopterus schreibersi Did Not Vanish. Journal of Israel Cave Research 1990: 97-98.
Italy
Aguzzi, Stefano, Bogliani, Giuseppe, Orioli, Valerio and Pilon, Nicola. (2017). Nehalennia speciosa rediscovered in northwestern Italy (Odonata: Coenagrionidae). Notulae odonatologicae 8(9): 319-374.
Baldini, Riccardo M. (1999). Fumana scoparia Pomel (Cistaceae): considerations on a poorly known species for the Italian flora / Fumana scoparla Pomel (Cistaceae), considerazioni su di una non ben nota specie della Flora Italiana. Webbia 54(1): 73-84.
Bianco P, Medagli P, D'Emerico S. 1989 Nuovi dati distributivi e osservazioni morfologiche su Aegilops uniaristata Vis. (Gramineae), entita mediterraneo-oriental riaccertata per la flora italiana. (Morphological observations on Aegilops uniaristata Vis. (Gramineae), eastern-Mediterranean entity rediscovered for the Italian flora.) Webbia 43. (1): 19-24.
Bonini, Federica et al. (2020). Juncus atratus Krock. (Juncaceae) rediscovered in Italy: a species deserving urgent conservation actions. Biologia. DOI: https://doi.org/10.2478/s11756-020-00490-2 [Abstract]
Canovi N., Gheza G., Pavesi M. & Villa D. 2014. Un nuovo importante sito di Sympecma paedisca (Brauer, 1877) (Odonata: Zygoptera: Lestidae) in Lombardia (Italia Settentrionale). Il Naturalista Valtellinese - Atti del Museo civico di Storia naturale di Morbegno, 25: 5–14.
Dondini, Gianna, Tomassini, A., Inguscio, S., and Rossi, E. (2014). Rediscovery of Mehely's horseshoe bat (Rhinolophus mehelyi) in peninsular Italy. Hystrix 25(1): 59-60.
Efetov, K. A.; Kucherenko, E. E.; Tarmann, G. M. (2020). An application of the synthetic sex attractants from the series "EFETOV-2" for studying Procridinae in Italy (Lepidoptera: Zygaenidae). Revista de Lepidopterologia 48(192): 733-749. [Abstract] [Rhagades pruni]
Fanelli, G., De Sanctis, M. and Serafini Sauli, A. (2012). La riscoperta di Trifolium latinum (Fabaceae) in Roma un secolo dopo la sua apparente sparizione dall’Italia. Informatore Botanico Italiano 44: 337-339.
Guariento, Elia and Franzini, Gabriele. (2021). Attalus minimus (Insecta, Melyridae) recently rediscovered in South Tyrol after 109 years. Gredleriana 21: 169-171.
Lorenzini, R., Lovari, S. and Masseti, M. (2002). The rediscovery of the Italian roe deer: Genetic differentiation and management implications. Italian Journal of Zoology 69: 367-379. [rediscovery in southern Tuscany]
Mendicino, Federica et al. (2026). New Records of Wild Bees from Calabria and BasilicataHighlight the Hidden Diversity of Anthophila in Italy. Diversity 18(2): 74. https://doi.org/10.3390/d18020074 [Andrena kamarti]
Mercurio, Vincenzo and Li Vigni, Fabrizio. (2007). Rediscovery of Pelobates fuscus insubricus in the Asti Province, north-western Italy. Acta Herpetologica 2: 1-6.
Piazzini, Sandro and Favilli, Leonardo. (2020). Rediscovery of a species considered to be extinct in the Pollino massif (Calabria, Italy): Erebia gorge (Hübner, [1804]) (Lepidoptera: Nymphalidae). Fragmenta Entomologica 52(1): 43-46.
Spinelli, Andrea et al. (2018). Returning of Hippocampus hippocampus(Linnaeus, 1758) (Syngnathidae) in the Faro Lake – oriented Natural Reserve of Capo Peloro, Italy. Natural Product Research. doi: https://doi.org/10.1080/14786419.2018.1490909 [Abstract]
Stinca, Adriano. (2017). Orchis italica Poir. (Orchidaceae): rediscovery after four centuries of a presumably extinct species in Mt. Vesuvius, Italy. Annali Di Botanica 7: 1-4.
Stinca, Adriano. (2023). Vegetation features of two vascular plant species presumed extinct and recently rediscovered in the natural habitat of community interest 8320 from Mt. Vesuvius, Italy. Plant Sociology 60(1): 13-23. doi:10.3897/pls2023601/02
Surdo, Salvatore. (2017). First record of Lindenia tetraphylla (Vander Linden, 1825) and rediscovery of Orthetrum nitidinerve (Selys, 1841) in Sicily (Insecta: Odonata). Fragmenta entomologica 49(2): 185-189.
Tavolara
Jamaica
"The Spotted Rail (Pardirallus maculatus) was presumed extinct in Jamaica for most of the period between its discovery by W.T. March in 1863 and its rediscovery in 1977 in the Upper Morass of the Black River system, St. Elizabeth Parish."
Japan
Akiyama, Yoshihiro B. and Yuhara, Takeshi. (2022). Rediscovery of the threatened gastropod Batillaria multiformis (Batillariidae) on the intertidal flat in the Edogawa Drainage Canal 27 years after the last observation. Biogeography 24: 53-63.
Hisashi ASHIDA and Kenji KITAYAMA, 1998. Rediscovery of Callytron inspeculare from Hyogo Prefecture. Ent. Rev. Japan, Osaka 53(1): 15-16.
Degawa, Yousuke et al. (2015). Rediscovery of Roesleria subterranea from Japan with a discussion of its infraspecific relationships detected using molecular analysis. MycoKeys 9: 1-9.
Iketani, Hiroyuki and Mase, Nobuko. (2015). Collection of Wild and Cultivated Rare Malus Genetic Resources in Northern Kyūshū, Japan. AREIPGR 31: 53-59.
Imahara, Y. and Ogawa, K. (2006). Rediscovery of Virgularia juncea (Octocorallia, Pennatulacea) from a tidal marsh in Okinawa, with a short note on its peculiar behavior, pp. 9-13. Proceedings of 10th International Coral Reef Symposium, Okinawa, Japan. June 28-July 2, 2004.
Imai, T., T. Oonuki, K. Matsuyama-Serisawa and Y. Serisawa. (2017). Rediscovery of freshwater prawn Macrobrachium formosense (Decapoda, Palaemonidae) ectoparasitized by green alga Cladogonium sp. from Shimama River, Tanega-shima Island, Kagoshima Prefecture, southern Japan. Nature of Kagoshima 43: 305-310.
Jørgensen, P. M. (2000). Survey of the lichen family Pannariaceae on the American continent, north of Mexico. The Bryologist 103: 670-704. [Fuscopannaria leucosticta]
Kato, Makoto and Sota, Teiji. (2022). Rediscovery of Macroplea japana (Coleoptera: Chrysomelidae: Donaciinae), an aquatic leaf beetle once thought to be extinct in Japan. Entomological Science 25(3): e12517. https://doi.org/10.1111/ens.12517
Kato, S., Higuchi, S., Kondo, Y., Kitano, S., Nozaki, H. and Tanaka, J. (2005). Rediscovery of the wild-extinct species Nitellopsis obtusa (Charales) in Lake Kawaguchi, Japan. J. Jpn. Bot. 80: 84-91.
Nagasawa, Kazuya and Nitta, Masato. (2015). Rediscovery of a Fish Acanthocephalan, Acanthocephalus minor (Echinorhynchida: Echinorhynchidae), in the Lake Biwa Basin, Central Japan, with a Review of the Fish Acanthocephalan Fauna of the Basin. Species Diversity 20: 73-81.
Nagasawa K., Yamamoto M., Sakurai Y. and Kumagai A. (1995). Rediscovery in Japan and host association of Salmincola carpionis (Copepoda: Lernaeopodidae), a parasite of wild and reared freshwater salmonids. Can. J. Fish. Aquat. Sci. 52: 178-185.
Naiki, Akiyo et al. (2023). Rediscovery of Exallage auricularia (Rubiaceae) from Yonaguni Island, Okinawa, Japan. The Journal of Japanese Botany 98(3): 134-139. https://doi.org/10.51033/jjapbot.jjapbot.ID0115
Nakajima, Hiroki and Ariyama, Hiroyuki. (2022). Record of Cloridopsis scorpio (Latreille in Latreille, Le Peletier, Serville & Guérin, 1828) (Stomatopoda: Squillidae) from Japan: rediscovery after almost a century. Plankton Benthos Res 17(2): 185-191. doi:10.3800/pbr.17.185
Okuno, Junji and Osawa, Masayuki. (2016). Rediscovery of Chirostylus dolichopus Ortmann, 1892 (Crustacea: Decapoda: Anomura: Chirostylidae) from its type locality, Boso Peninsula, Japan, with description of the colouration in life. Marine Biodiversity Records 9: 28.
Onuki, Keisuke and Fuke, Yusuke. (2022). Rediscovery of a native freshwater shrimp, Neocaridina denticulata, and expansion of an invasive species in and around Lake Biwa, Japan: genetic and morphological approach. Conservation Genetics 23: 967-980. https://doi.org/10.1007/s10592-022-01467-1
Sasaki, Mizuki, Iwaki, Takashi and Nakao, Minoru. (2022). Rediscovery of Michajlovia turdi (Digenea: Brachylaimoidea) from Japan. J. Parasitol. 108(2): 122-126. https://doi.org/10.1645/21-93
Tashiro, Yushu. (2022). Creating a Space for Creativity: Agricultural Canals and the Endangered Species Hemigrammocypris rasborella, pp. 51-65. In: Miyauchi, Taisuke and Fukunaga, Mayumi (eds.). Adaptive Participatory Environmental Governance in Japan: Local Experiences, Global Lessons. Springer Singapore. https://doi.org/10.1007/978-981-16-2509-1_4
Uchida, Kei et al. (2016). Rediscovery of Celes akitanus (Orthoptera: Acrididae) from semi-natural grasslands in Japan. Entomological Science 19(2): 89-96. [Abstract]
Watanabe, Reiya, Iwata, Yasuyuki and Kato, Atsushi. (2022). Rediscovery of Helophorus auriculatus Sharp, 1884 (Coleoptera, Helophoridae) in Saitama Prefecture, Japan, with ecological notes. 昆蟲(ニューシリーズ)25(3): 111-116.
Watanabe, Reiya and Ohba, Shin-ya. (2021). Rediscovery of Helophorus (Gephelophorus) auriculatus Sharp, 1884 (Coleoptera, Helophoridae) from Tsushima Island and Comparison of its Habitat, Morphology, and Mitochondrial COI among Another Japanese Population. Elytra, Tokyo, New Series 11(2): 295-300.
Yamana, Yusuke and Kohtsuka, Hisanori. (2016). Rediscovery of Plesiocolochirus inornatus (Marenzeller, 1881) (Echinodermata: Holothuroidea: Dendrochirotida: Cucumariidae) in Japan and new information on its external and ossicle morphology. Plankton & Benthos Research 11(2): 57-70.
Kenya
Korean Peninsula
Byun, Bong-Kyu. (In Press, 2018). Rediscovery of the little-known species of the subfamily Olethreutinae (Lepidoptera, Tortricidae) in the Korean Peninsula. Journal of Asia-Pacific Biodiversity. [three species: Olethreutes castaneana, Rudisociaria velutina and Phaecasiophora fernaldana]
Laos
Hämäläinen, Matti. (2020). A memorable encounter with Platycnemis phasmovolans, the ‘Flying Phantom Featherleg’ in Laos. Agrion 24(2): 84-88.
Latvia
Balalaikins, Maksims et al. (2018). New record of Calosoma (Campalita) auropunctatum (Herbst, 1784) in Latvia with notes on its occurrence in the Eastern Baltic region. Periodicum Biologorum 120(1): 59-65.
Jurciņš, Didzis et al. (2014). Refound of extinct lichen Lobaria amplissima (Scop.) Forssell in Latvia. Acta Biol. Univ. Daugavp. 14(1): 59-65.
Lebanon
Bury, Jaroslaw. (2014). Rediscovery of Charaxes jasius jasius (LINNAEUS, 1767) (Lepidoptera: Nymphalidae) in Lebanon. CESA News 97: 13-19.
Lesser Antilles
Martinique
Gros-Desormeaux, J.-R., T. Lesales, and A.-G. Tayalay. 2015. Behavioral observations on the White-breasted Thrasher (Ramphocinclus brachyurus brachyurus): conservation implications. Acta Ethologica 18: 197-208. [A pair was found in 1951 on the Caravelle Peninsula; cites Pinchon and Bon Saint Come, 1951]
Montserrat
Coote, Thomas, Schmidt, Robert E. and Schmidt, Kathleen A. (2021). Rediscovery of Populations of Pyrgophorus parvulus (Guilding, 1828) (Gastropoda, Cochliopidae), from a Freshwater Stream on Montserrat, West Indies. American Malacological Bulletin 38(2): 1-4. [Abstract]
Larsen, Roxanne et al. (2007). Mist netting bias, species accumulation curves, and the rediscovery of two bats on Montserrat (Lesser Antilles). Acta Chiropterologica 9(2): 423-435.
Libya
Meininger, P. .L, Wolf, P. A., Hadoud, D. A. and Essghaier, M. F. A. (1994). Rediscovery of Lesser Crested Terns breeding in Libya. British Birds 87: 160-170.
Lithuania
Gudžinskas, Zigmantas and Taura, Laurynas. (2022). Rediscovery of endangered species Laphangium luteoalbum (Asteraceae) in Lithuania. Botanica 28(1): 60-66. https://doi.org/10.35513/Botlit.2022.1.7
Macedonia
Jovanovska, Daniela et al. (2017). Rediscovery of Aldrovanda vesiculosa L. and new data on its distribution in the Republic of Macedonia. Acta Musei Macedonici Scientiarum Naturalium 20: 31-40.
Nahirnić-Beshkova A., Beshkov S., Kucherenko E.E. & Efetov K.A. (2021). Theresimima ampellophaga (Bayle-Barelle, 1808) rediscovered in the Republic of North Macedonia by using sex attractant traps (Lepidoptera: Zygaenidae). SHILAP Revista de Lepidopterología, 49(193): 161-170. https://doi.org/10.57065/shilap.332
Teofilovski, Aco. (2017). Contribution to knowledge of the flora of the Republic of Macedonia. Botanica Serbica 41(1): 99-103. [Lathyrus linifolius, Lactuca plumieri, Serratula tinctoria]
Madeiran Archipelago
Cameron, Robert A. D., Teixeira, Dinarte, Pokryszko, Beata, Silva, Isamberto and Groh, Klaus. (2021). An annotated checklist of the extant and Quaternary land molluscs of the Desertas Islands, Madeiran Archipelago. Journal of Conchology 44(1): 53-70. [Actinella actinophora; Deserta Grande (Desertas Islands)]
Medina, Leopoldo et al. (2021). Rediscovering of Chara braunii (Characeae, Charophyta) in Madeira (Macaronesian region, Portugal). Botanica Complutensis 45: e79754. https://doi.org/10.5209/bocm.79754
Malaya
Ali, S. and Santapau, H. (1958). Rediscovery of the smaller Asiatic one-horned rhinoceros (Rhinoceros sondaicus Desmarest) in Malaya. Jour. Bombay Nat. Hist. Soc. 55: 554-556.
Malaysia
HAMZA, A., MAMAT, I.B.H. & ABDULLAH, MT. 2019. Results of a seabird survey at the southern Seribuat Archipelago, Johor, Malaysia. Marine Ornithology 47: 49–53.
HAMZA, A. A., WONG, C.H. & AHMAD, A. 2016. Rediscovery of least known breeding sites for seabirds in East Coast Peninsular Malaysia. Malayan Nature Journal 68: 121-129.
Maideen, Haja et al. (2019). Rediscovery of Diplazium procumbens (Athyriaceae) in Peninsular Malaysia after more than 70 years, with notes on the morphology, spore ornamentation and anatomy. Phytotaxa 422(3): 241-247. [Abstract]
Quah, Evan S. H. et al. (2018). Identification and a new record from Penang Island of the rare redbellied reed snake (Calamaria albiventer) (Gray, 1835) (Serpentes: Calamariinae). Raffles Bulletin of Zoology 66: 486-493.
Sabah
Sarawak
Dinets, V. (2003). First photo of a bay cat in the wild? Cat News 38: 14.
Ramji, Mohamad Fizl Sidq et al. (2012). Rediscovery of the Enigmatic Mountain Blackeye, Chlorocharis emiliae Sharpe, 1888 (Passeriformes: Zosteropidae)from Mount Pueh, Sarawak. Tropical Natural History 12(2): 261-266. [Chlorocharis emiliae moultoni]
Malta
Lalov, S. V. (2007). Rediscovery of Asparagus stipularis Forskal (Family: Asparagaceae) in the Maltese Islands (Central Mediterranean). The Central Mediterranean Naturalist 4: 199-201.
Lalov, S. V. et al. (2007). Rediscovery of Ornithogalum divergens Boreau (Hyacinthaceae) in Malta (central Mediterranean). The Central Mediterranean Naturalist 4(3): 205-206.
Lanfranco, E. and Stevens, D. T. (2000). Rediscovery of Lotus halophilus Boissier et Spruner (Fabaceae) from the Island of Comino (Malta, Central Mediterranean). The Central Mediterranean Naturalist 3(2): 59-60.
Mifsud, Stephen. (2017). A Preliminary Survey and Taxonomy of Wild Roses (Rosa Spp.) Occurring on the Maltese Islands. Xjenza Online 5: 36-56. [Rosa canina, Rosa gallica]
Mifsud, Stephen, Napier, M., Fenech, S., and Cassar, L. F. (2016). Current status of Asplenium sagittatum (Aspleniaceae) in the Maltese islands. Fl. Medit. 26: 69-80.
Mariana Islands
Lemke, Dr. Thomas O. (1987). Sheath-tailed Bats Rediscovered in the Mariana Islands. BATS Magazine 5(1): (pagination?).
Richmond, Jonathan Q. et al. (2022). Range eclipse leads to tenuous survival of a rare lizard species on a barrier atoll. Oryx 56(1): 63-72. https://doi.org/10.1017/S0030605320001404
Mauritania
Shine, T., Böhme, W., Nickel, H., Thies, D. F. and Wilms, T. (2001). Rediscovery of relict populations of the Nile crocodile Crocodylus niloticus in south-eastern Mauritania, with observations on their natural history. Oryx 35: 260–262. [Abstract]
Mauritius
Florens, F. B. Vincent and Baider, Claudia. (2006). Relocation of ‘extinct’ Ficus densifolia Miq. (Moraceae) in Mauritius. Phelsuma 14: 101-103.
Mexico
Bean, Preston T. et al. (2010). Rediscovery of Headwater Catfish, Ictalurus lupus (Ictaluridae), in a Western Gulf Slope Drainage. DFC Annual Meeting 2010 Program.
Caviedes-Solis, I. W., L. F. Vázquez-Vega, I. Solano-Savaleta, E. Pérez-Ramos, S. M. Rovito, T. J. Devitt, P. Heimes, O. A. Flores-Villela, J. A. Campbell, and A. Nieto-Montes de Oca. 2015. Everything is not lost: recent records, rediscoveries, and range extensions of Mexican hylid frogs. Mesoamerican Herpetology 2: 230-241. [Agalychnis moreletti]
Delia, J. R. J., J. L. Whitney, and T. Burkhardt. 2013. Rediscovery of ‘lost’ treefrogs from Oaxacan highlands of Mexico. Biodiversity Conservation 22: 1,405–1,414. [Agalychnis moreletti]
Harper, Alan et al. (2016). Rediscovery of a high-altitude vole, Microtus californicus, in Baja California, Mexico. The Southwestern Naturalist 61(4): 333-338.
Janeba, Zlatko and Elizondo, Cecilia. (2022). Genus Mammillaria Haw. in the Yucatán Peninsula. Bradleya 2022(sp40): 150-160. https://doi.org/10.25223/brad.sp40.2022.a14 [Mammillaria columbiana yucatanensis]
Lagunas-Calvo, O., Hernández-Mena, D.I., Rivas, G., Contreraas-Mirón, S., Mendoza-Garfias, B., Echeverría-Martínez, O. M. and Oceguera-Figueroa, A. (2018). Rediscovery of the nemertean Prostoma graecense from Xochimilco, 75 years after its first and only record. Revista Mexicana de Biodiversidad 89: 39-45.
López-García, Margarita M., Deloya, C. and Gasca-Álvarez, H. J. (2016). Rediscovery of Tomarus cuniculus (Fabricius, 1801) (Coleoptera: Scarabaeidae: Dynastinae) in Mexico. Annales de la Société entomologique de France, New Series, 51: 494-498. [Abstract]
Nolasco-Luna, Jose Rafael, Cupul-Magaña, F. G., Escobedo-Galván, A. H., MataSilva, V. and Wilson, L. D. (2019). Rediscovery of Tantilla bocourti (Günther, 1895) on Isla María Cleofas, Nayarit, Mexico (Squamata: Colubridae). Herpetology Notes 12: 343-346.
O'Brien, Bart C., Rebman, Jon P., Delgadillo-Rodríguez, José, Gandhi, Kanchi Natarajan, Junak, Steven A., Oberbauer, Thomas A., Riemann, Hugo and Vanderplank, Sula E. (2025). The Rare, Endangered and Endemic Plants of the California Floristic Province Portion of Baja California, Mexico. Aliso 43(2): 77-217. https://doi.org/10.5642/aliso.YOAJ6071 [Downingia cuspidata in 2017 by James Riley; Allophyllum gilioides gilioides on Isla Guadalupe]
Sigel, Erin M. et al. (2014). Rediscovery of Polypodium calirhiza (Polypodiaceae) in Mexico. Brittonia 66: 278-286.
Solano-Gómez Rodolfo, Jiménez-Machorro, Rolando and Damon, Anne A. (2011). Two new records and one rediscovery for Orchidaceae of México. Acta Botanica Mexicana 96: 59-72. [Plectrophora alata]
Vázquez-Vega, L. F., I. W. Caviedes-Solis, I. Solano-Zavelata, R. Villegas-García, and O. A. Flores-Villela. 2015. Distribution Notes. Agalychnis moreletii (Duméril, 1853). Mesoamerican Herpetology 2: 538–539.
Guerrero
Padilla-Serrato, J. G., Soriano-Honorato, L. D., Kuk-Dzul, J. G., Flores-Garza, R., Torreblanca-Ramírez, C. and Campos, E. (2024). Rediscovery of Mesotheres unguifalcula (Glassell, 1936) (Crustacea: Brachyura: Pinnotheridae) with remarks on the symbiotic relationship with its new host, the spindle sea snail Leucozonia cerata (W. Wood, 1828) (Mollusa: Gastropoda: Fasciolariidae). Zool Stud 63:44. doi:10.6620/ZS.2024.63-44.
Moldova
Munjiu, Oxana. (2017). Distribution of endangered mayfly Palingenia longicauda (Olivier, 1791) (Ephemeroptera, Palingeniidae) on the territory of the Republic of Moldova. Lauterbornia 84: 39-51.
Mongolia
Gundegmaa, Vanjil et al. (2022). The vascular flora of the Sutai Khairkhan Mountain Nature Reserve, Mongolia. Botanica Pacifica. A journal of plant science and conservation 11(1): 115-128. doi:10.17581/bp.2022.11102 [Microula tibetica pratensis]
Morocco
El Alami, Abderrazak, Bouzid, El Mustapha and Fattah, Aderrazzak. (2022). Rediscovery of the striped hyaena Hyaena hyaena in the central High Atlas after 22 years. Oryx 56(5): 650. https://doi.org/10.1017/S003060532200076X
Escoriza, D., Comas, M. M., Donaire, D. and Carranza, S. (2006). Rediscovery of Salamandra algira Bedriaga, 1883 from the Beni Snassen massif (Morocco) and phylogenetic relationships of North African Salamandra. Amphibia-Reptilia 27(3): 448-455.
Myanmar
Oo, Sai Sein Lin and Bates, Paul Jeremy James. (2016). The rediscovery of the common water monitor lizard Varanus salvator (Squamata: Varanidae) in northern Myanmar. Journal of Threatened Taxa 8(5): 8827-8828.
Nepal
Raju Acharya, Yadav Ghimirey, Geraldine Werhahn, Naresh Kusi, Bidhan Adhikary, Binod Kunwar. (2015). Wild yak Bos mutus in Nepal: rediscovery of a flagship species. Mammalia. DOI: 10.1515/mammalia-2015-0066 [Abstract]
Netherlands
Kooijman, Annemieke, Hedenäs, Lars, Mettrop, Ivan and Cusell, Casper. (2015). Calliergon megalophyllum rediscovered in the Netherlands after 50 years: comparison to Swedish habitats. Lindbergia 38: 20-29.
Sanabria, M. J., Lock, K., Scheepens, M. and Tempelman, D. (2012). Oligostomis reticulata (Linnaeus, 1761) (Trichoptera, Phryganeidae) recorded again in The Netherlands and Belgium. Lauterbornia 75: 1-13.
van den Berg LJ, te Linde BH. 2001 Brede wolfsmelk (Euphorbia platyphyllos L.) weer gevonden in het oostelijk rivierengebied. (Euphorbia platyphyllos L. rediscovered in the Nederlands.) Gorteria 27. (1): 1-4.
Nehallennia speciosa
New Caledonia
BARGIER, N., DOMINIQUE, Y., UMBERT, J.-F., PARPET, J.-F., LACROIX, G., JEANPERT, J. & GENTON, P. (2018). Diagnose des Dolines de Nouvelle-Calédonie. – [Rapport de recherche] CNRT “Nickel et son environnement”, 136 pp. https://hal.archives-ouvertes.fr/hal-02152037 [Phalilus oberthuri]
Pignal, Marc, Laudereau, Christian, Laudereau, Pierre.-Louis and Bruy, David. (2022). Une révision de Dendrobium Sw. sect. Rhizobium Lindl. en Nouvelle-Calédonie. Adansonia, sér. 3, 44(15): 153-164. https://doi.org/10.5252/adansonia2022v44a15
van Vondel, B. J. et al. (2021). On the rediscovery of Phalilus oberthuri (GUIGNOT, 1935) in New Caledonia (Coleoptera: Haliplidae). Koleopterologische Rundschau 91: 311-322. [Phalilus oberthuri]
New Guinea
Bednarek-Ochyra, Halina and Plášek, Vítězslav. (2017). Occurrence of Racomitrium pruinosum (Grimmiaceae, Bryophyta) in New Guinea, with a Review of Gondwanan Mosses in the Tropics. Herzogia 30(2): 412-426. [Abstract]
New Zealand
Cameron EK. 2000 Native sow thistle, Sonchus kirkii, rediscovered in the Auckland region. Auckland Bot. Soc. J. 55. (1): 21-24.
Laing, Robert M. (1914). A Revised List of the Norfolk Island Flora, with some Notes on the Species. Transactions of the New Zealand Institute 47: 1-39. [Lycopodium drummondii]
Mary Morgan-Richards, M. Rheyda Hinlo, Chris Smuts-Kennedy, John Innes, Weihong Ji, Manuela Barry, Dianne Brunton, and Rodney A. Hitchmough. (In Press, 2015). Identification of a Rare Gecko from North Island New Zealand, and Genetic Assessment of Its Probable Origin: A Novel Mainland Conservation Priority? Journal of Herpetology. [Abstract]
Niue
Norway
Flåten, M. and Fjellberg, A. (2008). Rediscovery of Osmoderma eremita (Scopoli, 1763) (Coleoptera, Scarabaeidae) in Norway. Norwegian Journal of Entomology 55: 165-168.
Kjærandsen, Jostein. (2022). Rocetelion humerale (Zetterstedt, 1850) (Diptera, Keroplatidae) rediscovered in Norway after more than 100 years, with description of the larva and its habitat. Norwegian Journal of Entomology 69, 000–000.
Pakistan
Ahmed, Quratulan and Ali, Qadeer Mohammad. (2020). Holothurians from Pakistan: New addition of Holothuria (Theelothuria) notabilis (Ludwig, 1875) and rediscovery of Actinocucumis typica (Ludwig, 1875) from the Karachi coast, northern Arabian Sea. SPC Beche-de-mer Information Bulletin 40: 40-42.
Awan, Muhammad Naeem et al. (2019). Rediscovery and first nesting record of the Vulnerable Cheer Pheasant Catreus wallichii in Machiara National Park, Kashmir Himalaya, Pakistan. BirdingASIA 31: 79-84.
Baig, K. J. (2002). Rediscovery of Murree Hill Frog, Paa vicina after 130 years from Ayubia National Park. Proceedings of Pakistan Academy of Sciences 39(2): 261-262.
Mebert, Konrad et al. (2013). The Dice Snake, Natrix tessellata (Serpentes: Colubridae) in Pakistan: Analysis of its Range Limited to Few Valleys in the Western Karakoram. Pakistan J. Zool. 45(2): 395-410.
Mebert, Konrad and Masroor, Rafaqat. (2013). Dice Snakes in the western Himalayas: discussion of potential expansion routes of Natrix tessellata after its rediscovery in Pakistan. Salamandra 49(4): 229-233. [automatic download]
Morandini, André C. and Gul, Shahnawaz. (2016). Rediscovery of Sanderia malayensis and remarks on Rhopilema nomadica record in Pakistan (Cnidaria: Scyphozoa). Papéis Avulsos de Zoologia 56(15): 171‑175.
Roberts, T. J. (1999). Rediscovery of the Afghan Mole Ellobius fuscocapillus in Pakistan. Journal of the Bombay Natural History Society 96: 134-139.
Shah, B., Hassan, M.A., Naveed, H., Shakeel, M., Khan, M.T. & Duan, Y.N. (2023) New findings of Deltocephalinae (Hemiptera, Cicadellidae) from Pakistan. Journal of Insect Biodiversity and Systematics 9(3): 583-590. https://doi.org/10.52547/jibs.9.3.583 [Platymetopius fidelis]
Tirmizi, N. M. and Ghani, N. (1994). Recovery of a Leucosid crab Nursia abbreviata Bell, 1855 from Arabian sea fish. Pakistan J. of Mar. Sci. 3(1): 83-85.
Papua New Guinea
Keim, A. P. (2009). Pandanaceae of the island of Yapen, Papua (West New Guinea), Indonesia, with their nomenclature and notes on the rediscovery of Sararanga sinuosa, and several new species and records. Blumea 54: 255-266.
White, W. T., Appleyard, S. A., Sabub, B., Kyne, P. M., Harris, M., Lis, R. et al. (2015). Rediscovery of the Threatened River Sharks, Glyphis garricki and G. glyphis, in Papua New Guinea. PLoS ONE 10(10): e0140075.
Paraguay
Satterlee, S. Andrew et al. (2012). Rediscovery of Aeglid Crabs in the Rio Jejuí Watershed, Paraguay. Journal of Crustacean Biology 32(4): 541-543. [Abstract]
Perú
Quiñones, Javier et al. (2014). Rediscovery of the Yellow-bellied Sea Snake, Hydrophis platurus (Linnaeus, 1766) in Máncora, northern Perú. Check List 10(6): 1563-1564. [automatic download]
Philippines
Allen, D. (2009). Worcester's Buttonquail Turnix worcesteri rediscovered at Dalton Pass. BirdingASIA 11: 83-84. [Luzon, Philippines]
Beukema, Wouter. (2011). First record of the genus Tropidonophis (Serpentes: Colubridae) and rediscovery of Parias flavomaculatus (Serpentes: Viperidae) on Siquijor Island, Philippines. Herpetology Notes 4: 177-179.
Tan, Benito C. et al. (2017). Additions to the moss floras of Mindanao and the Philippines with a focus on the rediscovery of Euptychium setigerum. Nat. Hist. Bull. Siam Soc. 62(1): 15-20.
Biag, Rachel D. and Alejandro, Grecebio Jonathan D. (2020). Rediscovery of Psychotria species, subspecies, and varieties collected in the 1990s and new records of Antirhea benguetensis (Elmer) Valeton and Ixora longifolia Smith (Rubiaceae) in Northern Sierra Madre Natural Park, Luzon, Philippines. Biodiversitas 21(10): 4524-4535.
GONZALEZ JC, DANS ATL, PEDREGOSA MG, CHIU C, VILLAHERMOSA R. 1999. Island-wide survey of forest and fauna and flora inventory of selected sites for priority conservation on Cebu. Manila: Fauna & Flora International (unpublished). [Copsychus cebuensis]
van de Ven, Willem et al. (2017). Movements and home ranges of Philippine crocodiles in San Mariano, lsabela. NMP Journal of Natural History 2(1): 38-45.
Gallicolumba luzonica (Polillo population)
Poland
Blaik T., Malkiewicz A. & Wasala R. 2011: Rediscovery and remarks on occurrence of Thaumetopoea processionea (Linnaeus, 1758) (Lepidoptera: Notodontidae: Thaumetopoeinae) in Poland. Wiadomości Entomologiczne
30(4): 246-256.
Brzeska, Paulina et al. (In Press, 2015). New records of Chara connivens P. Salzmann ex A. Braun 1835 – an extremely rare and protected species in Polish brackish waters. Acta Societatis Botanicorum Poloniae. DOI: 10.5586/asbp.2015.010.
Bury, J. (2020). Common Glider – Neptis sappho (Pallas, 1771) (Lepidoptera: Nymphalidae) in south-eastern Poland: another case of oversight or rapid range expansion of a species considered extinct? ANNALS OF THE UPPER SILESIAN MUSEUM IN BYTOM ENTOMOLOGY Vol. 29 (online 006): 1-20.
Górski, P. and Romański, M. (2016). Rediscovery of the rare liverwort Liochlaena subulata (Jungermanniaceae) in Poland. Herzogia 29(2): 810–813. [Abstract]
Gruzik, Janusz and Paul, Wojciech. (2000). Veronica peregrina (Scrophulariaceae) in Kraków – rediscovered after a century. Fragm. Flor. Geobot. 45(1-2): 513–539.
Guzik, Piotr and Guzik, Wojciech. (2022). Rediscovery of Isophya stysi (Čejchan, 1957) (Orthoptera: Tettigonioidea: Phaneropterinae) in south-eastern Poland. Fragmenta Faunistica 65(1): 69-75. doi:10.3161/00159301FF2022.65.1.069
Krajewski, Łukasz. (2017). Drepanocladus turgescens (Bryophyta, Amblystegiaceae) Rediscovered in Poland. Cryptogamie, Bryologie 38(3): 265-273. [Abstract]
Marciniuk P, Marciniuk J, Łysko A, Krajewski Ł, Chudecka J, Skrzyczyńska J, Popiela AA. 2020. Rediscovery of Cyperus flavescens (Cyperaceae) on the northeast periphery of its range in Europe. PeerJ 8:e9837 https://doi.org/10.7717/peerj.9837
Markowski, Piotr and Buczyński, Paweł. (2020). Rediscovery of Dytiscus latissimus Linnaeus, 1758 (Coleoptera: Dytiscidae) on the southern edge of its distribution area in Central and Eastern Poland. Polish Journal of Entomology 89(2): 81-90.
Plášek, Vítězslav, Číhal, Lukáš et al. (2024). Newly found and rediscovered hornworts (Anthocerotophyta) in Poland: Indicators of climate change impact in Central Europe. PhytoKeys 248: 237-261. https://doi.org/10.3897/phytokeys.248.134729
Plášek, Vítězslav, Ochyra, R., Smoczyk, M., Wiaderny, A. and Koopman, J. (2013). Recent rediscovery of the epiphytic moss Orthotrichum pulchellum Brunt. in Poland. Čas. Slez. Muz. Opava (A), 62: 97-100.
Plášek, Vítězslav et al. (2016). Rediscovery of the epiphytic moss Ulota coarctata (Orthotrichaceae) in Poland. Polish Botanical Journal 61(1): 99-105.
Stebel, Adam. (2010). Rediscovery of Orthotrichum rogeri Brid. (Bryophyta) in Poland. Opole Scientific Society Nature Journal 43: 23-27.
Szeląg, Zbigniew and Wójcik, Grzegorz. (In Press, 2014). Hieracium sudetubulosum (Asteraceae) rediscovered outside the Karkonosze Mts. Polish Botanical Journal 59(1): 1-xx. [automatic download]
Vončina, Grzegorz, Cykowska, Beata and Chachuła, Piotr. (2011). Rediscovery of Buxbaumia viridis (Bryophyta, Buxbaumiaceae) in the Tatraand Gorce in the Polish western Carpathians, pp. 171-176. In: Stebel, A. and Ochyra, R. (eds.). Chorological Studies on Polish Carpathian Bryophytes, Sorus, Poznań.
Szeląg, Zbigniew. (2011). Hieracium albinum (Asteraceae) rediscovered. Polish Botanical Journal 56: 77-79.
Portugal
Cunha, Alexander Henriques et al. (2013). The rediscovery of Caulerpa prolifera in Ria Formosa, Portugal, 60 years after the previous record. Notebooks Marine Biology 54(3): 359-364.
Sérgio, C., Rodríguez-González, P. M., Albuquerque. A. and Garcia, C. A. (2011). Rediscovery of Rhamphidium purpuratum Mitt. (Bryophyta, Ditrichaceae) in Portugal after more than 60 years. Field Bryology 105: 40-41.
Puerto Rico
Caraballo-Ortiz, Marcos A. (2013). Rediscovery of Arrabidaea chica (Bignoniaceae) and Entada polystachya var. polyphylla (Fabaceae) in Puerto Rico. Phytotaxa 125(1): 53-58. [both local rediscoveries]
Réunion (French)
Albert, S., Rhumeur, A., Rivière, J. L., Chauvrat, A., Sauroy-Toucouère, S., Martos, F. and Strasberg, D. (2017). Rediscovery of the mistletoe Bakerella hoyifolia subsp. bojeri (Loranthaceae) on Reunion Island: population status assessment for its conservation. Botany Letters 164(3): 229-236. https://doi.org/10.1080/23818107.2017.1340191
Romania
Balázs, Z. R., Roman, A., Balázs, H. E., Căpraș, D., Podar, D. (2016) Rediscovery of Cypripedium calceolus L. in the vicinity of Cluj-Napoca (Romania) after 80 years, Botanical Contribution, LI: 43-53.
Bartha, L. and Bartók, A. (2013). Rediscovery of Astragalus roemeri Simonk. in the Scăriţa-Belioara Natural Reserve (Apuseni Mountains, Romania). Contributii Botanice 48: 23-26. [Abstract]
Bobocea, M. M., Tomescu, C. V. and Moscaliuc, L. A. (2020). The rediscovery of the rare Carpathian endemic orchid Nigritella carpatica in Romania after 139 years. Ber. Arbeitskrs. Heim. Orchid. 37(2): 188-210.
Cazan, Cristina Daniela et al. (2021). Seasonal dynamics of Phlebotomus neglectus (Diptera: Psychodidae) in cave microhabitats in Romania and the rediscovery of Sergentomyia minuta (Rondani, 1843) after 50 years. Parasites & Vectors 14: 476. https://doi.org/10.1186/s13071-021-04985-y
Cserkész, T., Aczél‐Fridrich, Z., Hegyeli, Z., Sugár, S., Czabán, D., Horváth, O., Sramkó, G. (2015): Rediscovery of Hungarian birch mouse (Sicista subtilis trizona) in Transylvania (Romania) with molecular characterisation of its phylogenetic affinities. Mammalia 79: 215‐224.
Dincă, V. and Vila, R. (2008). Improving the knowledge on the Romanian Rhopalocera, including the rediscovery of Polyommatus amandus (Schneider, 1792) and an application of DNA-based identification. Nota Lepidopterologica 31(1): 3-23.
Laszlo, R., Cosmin Ovidiu, M., Claudia Maria, C. and Demetra, R. (2024). Rediscovered only to be Almost Exterminated Again - The Story of Tomares nogelii dobrogensis (Nogel’s Hairstreak) a Charismatic Butterfly Endemic to Europe. Ecol Conserv Sci. 4(3): 5556366. http://dx.doi.org/10.19080/ECOA.2024.04.555636
Mátis, Attila et al. (2014). Lost and found: rediscovery of Saussurea porcii Degen in the Rodnei Mountains (Eastern Carpathians, Romania) after more than a century. Contribuţii Botanice XLIX: 39-42.
Pintilioaie, Alexandru-Mihai and Urák, István. (2022). The rediscovery of Aculepeira armida (Araneae: Araneidae) in Romania after more than 100 years. Biharean Biologist 16(1): 40-43.
Rákosy, L. and Craioveanu, C. (2015). Redescovering Tomares nogelii dobrogensis Caradja, 1895 in Romania. Entomologica Romanica 19: 13-16.
Russia
Efimov, P. G., Legchenko, M. V. and Sheludyakova, M. B. (2025). Taraxacum norvegicum (Asteraceae) in Russia: historical records, rediscovery and issues of conservation (Taraxacum Notulae Rossiae Boreali Europeae V). Novitates Systematicae Plantarum Vascularium 56: e06. [Abstract]
Popov, Igor. (2021). In the search of the lost pearl: Rediscovery of Southern Populations of Margaritifera margaritifera (L.) in Russia as a Model of Conservation Research. Springer Nature. [Abstract]
Scotland
Cosgrove, Peter. (2002). The Cairngorms Local Biodiversity Action Plan. 200 pp. [Buxbaumia viridis]
Shetland Islands
Lugg, Keith et al. (2022). Re-discovery of Geophilus proximus C.L.Koch, 1847 in Shetland (Chilopoda: Geophilomorpha, Geophilidae). Bulletin of the British Myriapod & Isopod Group 34: 2-6.
Serbia
SLOBODAN IVKOVIĆ et al. (2020). Rediscovery of Southern Barbed-wire Bush-cricket, Onconotus servillei Fischer von Waldheim (Orthoptera: Tettigoniidae: Tettigoniinae: Onconotini) in Serbia, with notes on species’ calling song at the westernmost border of its distribution area. Zootaxa 4732(4): 596-600. [Abstract]
Steenis, Jeroen van et al. (2019). New records of hoverflies (Diptera: Syrphidae) and the rediscovery of Primocerioides regale Violovitsh for the fauna of Serbia. Biologia Serbica 41(1): 94-103.
Trbojević, Ivana et al. (2019). Rediscovery of Chara canescens Loiseleur in Serbia. Botanica Serbica 43(1): 97-102.
Vujić, Mihailo, Reljić, Kosta, Franeta, Filip and Tot, Ivan. (2026). Rendezvous in the woods: rediscovery of Lopinga achine (Scopoli, 1763) (Lepidoptera: Nymphalidae) in Serbia. Kragujevac J. Sci. 48 (1).
Seychelles
Krijn Buitelaar and Philip de Pous. (2011). First record of Pelusios castanoides intergularis and rediscovery of Pelusios subniger parietalis on Cousin Island, Seychelles. Herpetology Notes 4: 9-10.
Singapore
Ang, W. F., Lok, A. F. S. L. and Tan, H. T. W. (2010). Rediscovery in Singapore of Pinanga simplicifrons (Miq.) Becc. (Arecaceae). Nature in Singapore 3: 83-86.
Ang, W.F., Lok, A.F.S.L., Yeo, C.K., Angkasa, A., Ng, P.X. Ng, B.Y.Q. & Tan, H.T.W. (2011). Rediscovery of Renanthera elongata (Blume) Lindl. (Orchidaceae) in Singapore. Nat. Singapore 4: 297-301.
Ang, W.F., Lok, A.F.S.L., Yeo, C.K., Angkasa, A., Ng, P.X. & Tan, H.T.W. (2012). Rediscovery of Freycinetia javanica Blume (Pandanaceae) in Singapore. Nat. Singapore 5: 117-122.
Ang, W. F., Lok, A. F. S. L., Yeo, C. K., Tan, S. Y. and Tan, H. T. W. (2010). Rediscovery of Dendrobium aloifolium (Blume) Rchb. f. (Orchidaceae) in Singapore. Nature in Singapore 3: 321-325.
Ang, W.F., Yeo, C.K., Lok, A.F.S.L., Angkasa, A., Ng, P.X. & Tan, H.T.W. (2012). Rediscovery of Trichotosia velutina (Lodd. ex Lindl.) Kraenzl. (Orchidaceae) in Singapore. Nat. Singapore 5: 199-204.
Ang, W.F., Yeo, C.K., Ooi, Z.Y., Lok, A.F.S.L., Abdullah, I.H. & Tan, H.T.W. (2014). Rediscovery in Singapore of Ficus delosyce Corner (Moraceae). Nat. Singapore 7: 167–172.
Ascher, John S. et al. (2022). The bees of Singapore (Hymenoptera: Apoidea: Anthophila): First comprehensive country checklist and conservation assessment for a Southeast Asian bee fauna. Raffles Bulletin of Zoology 70: 39-64. [Thyreus abdominalis rostratus]
Chong KY, Lim RCJ, Loh JW, Neo L, Seah WW, Tan SY & Tan HTW. (2018). Rediscoveries, new records, and the floristic value of the Nee Soon freshwater swamp forest, Singapore. Gardens’ Bulletin Singapore, 70 (Supplement 1): 49-69. [Baccaurea macrophylla]
Choo, L. M. and Ngo, K. M. (2020). A revision of the genus Sindora (Fabaceae, Detarioideae) in Peninsular Malaysia and Singapore. Gardens' Bulletin Singapore 72(2): 233-254.
Chua, M., Sivasothi, N. & Teo, R., 2009. Rediscovery of the greater mousedeer Tragulus napu (Mammalia: Artiodactyla: Tragulidae) in Pulau Ubin, Singapore. Nature in Singapore 2: 373-378.
Ganesan, S. K. (2013). Status and distribution of Pterospermum species in Singapore. Nature In Singapore 6: 149-152. [P. diversifolium]
Ho, B. C. and Lua, H. K. (2022). Flora of Singapore precursors, 32: Discoveries in Mucuna (Leguminosae, subfamily Papilionoideae) with a review of the genus in Singapore. Gardens’ Bulletin Singapore 74(1): 37-55.
Ho, B. C. et al. (2018). New records and rediscoveries of vascular plants in Bukit Timah Nature Reserve, Singapore. Gardens' Bulletin Singapore 70(1): 33-55. [Aglaia palembanica, Bolbitis sinuata, Calamus ornatus, Claoxylon longifolium, Dapania racemosa, Dioscorea kingii, Ficus rosulata, Lasianthus reticulatus, Ryparosa hullettii, Senegalia kekapur]
Ibrahim, Ali et al. (2023). Biodiversity Record: Rediscovery of Engelhardia serrata in Singapore. Nature In Singapore 16: e2023047. doi:10.26107/NIS-2023-0047
Ibrahim, Hassan, Leong, P. K. F., Yam, T. W. and Lua, H. K. (2011). The rediscovery of the ground orchid Dienia ophrydis (J. König) Seidenf. (Orchidaceae) in Singapore. Nature in Singapore 4: 329-337.
Jain, Anuj et al. (2019). Rediscovery of the orange gull butterfly, Cepora iudith malaya, in Singapore. Singapore Biodiversity Records 2019: 22-23.
Lam WN, Loh JW, Chong R, Ting YY, Chan PJ, Rahman NE & Chong KY. (2022). Towards a field guide to the trees of the Nee Soon Swamp Forest (VII): Phyllanthaceae. Nature in Singapore, Supplement no. 1: e2022073. DOI: 10.26107/NIS-2022-0073 [Baccaurea pyriformis, Bridelia cinnamomea]
Lee, S. M. L. et al. (2021). Rediscovery of the Sculptured Toadstool, Amanita sculpta (Amanitaceae) in Singapore. Gardens' Bulletin Singapore 73(1): 1-7. doi: 10.26492/gbs73(1).2021-01
Leong, Paul K. F. et al. (2018). Acriopsis ridleyi Hook.f. (Orchidaceae): Re-encounter of an orchid thought extinct since its 1889 holotype collection in Singapore. Nature in Singapore 11: 27-36.
Leong, T.M. & Lim, K.K.P. (2009). Noteworthy microchiropteran records from the Bukit Timah and Central Catchment Nature Reserves, Singapore. Nat. Singapore 2: 83-90. [Bukit Timah Nature Reserve: Hipposideros bicolor]
Leong, P.K.F. & Yam, T.W. (2013). The rediscovery of Hetaeria obliqua in Singapore. Malesian Orchid J. 12: 23-28.
Leow, Calvin Jiah Jay. (2021). Biodiversity Record: A rediscovery and three new molluscan records for Singapore. Nature in Singapore 14: e2021110. DOI: 10.26107/NIS-2021-0110 [Swainsonia ocellata]
Lim, Norman T-L. and Xiuling, Ou Yang. (2012). Occurence of the Malay civet , Viverra tangalunga (Mammalia: Carnivora: Vivierridae) in Singapore. Nature In Singapore 5: 79-81.
Lim, Reuben C. J. et al. (2014). Rediscovery in Singapore of Vrydagyznea lancifolia Ridl. (Orchidaceae). Nature in Singapore 7: 1-7.
Lin, Jiayuan. (2022). Rediscovery of the thumbprint emperor, Lethrinus harak, in Singapore. Nature in Singapore 15: e2022148.
Loh, Jess et al. (2022). Biodiversity Record: Rediscovery of the butterfly, Rapala cowani, on Singapore Island. Nature in Singapore 15: e2022113.
Lok, A. F. S. L., W. F. Ang, K. Y. Chong & H. T. W. Tan. (2010). Rediscovery of Liparis barbata Lindl.(Orchidaceae) in Singapore. Nature in Singapore 3: 277-281.
Lok, A. F. S. L., W. F. Ang, K. Y. Chong, C. K. Yeo & H. T. W. Tan, 2011. Rediscovery in Singapore of Coelogyne rochussenii de Vriese (Orchidaceae). Nature in Singapore 4: 49-53.
Lok, A.F.S.L., Ang, W.F. & Yeo, C.K. (2012). Rediscovery of Callostylis pulchella (Lindl.) S.C.Chen & Z.H.Tsi (Orchidaceae) in Singapore. Nat. Singapore 5: 205-209.
Lok, A. F. S. L., K-x. Tan and H. T. W. Tan. (2008). Rediscovery of Dendrobium lobbii Teijsm. & Binnend. (Orchidaceae) in Singapore. Nature in Singapore 1: 35-39.
Lok, A.F.S.L., Ang, W.F. & Tan, H.T.W. (2011). The rediscovery in Singapore of Polystachya concreta (Jacq.) Garay & H.R. Sweet (Orchidaceae). Nat. Singapore 4: 19-24.
Lok, A. F. S. L. and Tan, H. T. W. (2008). Rediscovery of Aeschynanthus albidus (Blume) Steud. (Gesneriaceae) in Singapore. Nature in Singapore 1: 5-8.
Loo, Adrian H. B. (2011). Rediscovery in Singapore of Salacca affinis Griff. (Arecaceae). Nature in Singapore 4: 123-126.
Loo, Adrian H. B. et al. (2017). Rediscovery in Singapore of Calamus densiflorus Becc. (Arecaceae). Nature in Singapore 10: 1-5.
Ng, P.K.L. & Lim, K.K.P. (1989). Rediscovery of the Dwarf Snakehead, Channa gachua (Hamilton, 1822) (Channidae) in Singapore. Raffles B. Zool. 37(1): 172-174.
Niissalo, Matti A. (2023). Records of two extant orchid species (Orchidaceae) in Singapore. Nature in Singapore 16: e2023023. [Dendrobium singaporense, Bulbophyllum gusdorfii]
Niissalo, Matti A. et al. (2016). Rediscovery in Singapore of Plocoglottis lowii Rchb.f. (Orchidaceae). Nature in Singapore 9: 41-46.
Ong, Xin Rui, Chua, Siew Chin and Potts, Matthew D. (2013). Recent records of the dung beetle Catharsius molossus (Coleoptera: Scarabaeidae) in Singapore. Nature in Singapore 6: 1-6.
Ooi, Z. Y. and Ang, W. F. (2015). Rediscovery in Singapore of Lepironia articulata (Retz.) Domin (Cyperaceae). Nature in Singapore 8: 65-68.
Pottie, S.A. (1996). Studies of the Ecology and Behaviour of Insectivorous Bat Species in Singapore. Unpublished M.Sc. thesis, School of Biological Sciences, National University of Singapore. [Bukit Timah Nature Reserve: Emballonura monticola]
Rodda, Michele and Lai, Joseph. (2018). The rediscovery of Hoya obtusifolia (Apocynaceae, Asclepiadoideae) in Singapore. Nature in Singapore 11: 45-51.
Rodda, M., Tran, H. D., Chiew, P. T., Liew, D. and Leong-Škorničková, J. (2012). The rediscovery of Dischidia hirsuta (Apocynaceae, Asclepiadoideae) in Singapore. Gardens’ Bulletin Singapore 64(2): 293-299.
Serin, S., Law, I. S. and Thomas, N. (2017). Rediscovery of Gimlett’s reed snake in Singapore. Singapore Biodiversity Records 2017: 89-90.
Sheue, Chiou-Rong, Yong, Jean W. H. and Yang, Yeun-Po. (2005). The Bruguiera (Rhizophoraceae) Species in the Mangroves of Singapore, Especially on the New Record and the Rediscovery. Taiwania 50(4): 251-260. [Bruguiera sexangula]
Soh, Z. W. W. and Ngiam, R. W. J. (2013). Flower-visiting bees and wasps in Singapore parks (Insecta: Hymenoptera). Nature in Singapore 6: 153-172. [Megachile ramera]
Soh, Z. S.-H. and Ngiam, R. W. J (2025). Biodiversity Record: Damselflies Mortonagrion falcatum and Aciagrion borneense at Tuas. Nature in Singapore 18: e2025051. doi:10.26107/NIS-2025-0051 [Mortonagrion falcatum]
Tan, Siong Kiat et al. (2015). Terrestrial snails and slugs (Mollusca: Gastropoda) of Pulau Tekong, Singapore. Nature in Singapore 8: 25-30. [Paropeas achatinaceum]
Tan, S. K., Tan, S. H. and Low, Martyn E. Y. (2011). A reassessment of Verpa penis (Linnaeus, 1758) (Mollusca: Bivalvia: Clavagelloidea), a species presumed nationally extinct. Nature in Singapore 4: 5-8.
Tan, Tsu Soo and Toh, Chay Hoon. (2019). Rediscovery of spotfin frogfish in Singapore. Singapore Biodiversity Records 2019: 141-142.
Teo, R.C.H. & Rajathurai, S. (1997). Mammals, reptiles and amphibians in the nature reserves of Singapore – diversity, abundance and distribution. Proceedings of the Nature Reserves Survey Seminar. Gard. Bull. Singapore 49: 353–425. [Bukit Timah Nature Reserve: Pelophryne signata, Tylonycteris pachypusIomys horsfieldii]
Tsang, Kwok Choong. (2020). Rediscovery of the spotted guava blue butterfly in Singapore. Singapore Biodiversity Records 2020: 128. [Virachola subguttata malaya]
Williams, C. (2014). The rediscovery of a presumed nationally extinct Aeschynanthus. Gardenwise 43: 10-11. [Aeschynanthus pulcher, rediscovered in 2013 in Nee Soon Swamp Forest]
Yam, T.W., Leong, P.K.F., Chew, P.T., Liew, D. & Ng, W.K.H. (2010). The re-discovery and conservation of Bulbophyllum singaporeanum. Gardenwise 35: 15-17.
Yeo, C.K., Ang, W.F. & Lok, A.F.S.L. (2012). Pterisanthes (Vitaceae) of Singapore: with a note on the rediscovery of Pterisanthes cissioides Blume. Nat. Singapore 5: 185-190.
Yong, D.L., Fam, S.D. and Ng, J.J. (2008). Rediscovery of Dumeril's Monitor Varanus dumerilii (Varanidae) in Singapore. Nature in Singapore 1: 21-22.
Slovakia
Dítě, Daniel et al. (2017). Triglochin maritima – rediscovered in southern Slovakia. Thaiszia - J. Bot., Košice, 27(2): 129-136.
Dítě, D., Balla, M. and Dítě, Z. (2021). Ranunculus polyphyllus Willd. still grows in Slovakia. Thaiszia - J. Bot. 31 (2): 205-212.
Fuljer, Filip, Jamrich, Michal and Kautmanová, Ivona. (2023). Rediscovery of Lactarius nanus in Slovakia after 24 years. Acta Botanica Universitatis Comenianae 61: 37-50.
Hemala, V. et al. (2017). Additional records of Tropidothorax leucopterus (Goeze, 1778) (Hemiptera: Heteroptera: Lygaeidae) for Slovakia. Naturae Tutela 21(1): 51-58. [summarises previous rediscoveries]
Hemala, V. et al. (2018). Rediscovery of the rare flat bug Mezira tremulae (Hemiptera: Heteroptera: Aradidae) in Slovakia. Naturae Tutela 22(1): 69-78. [the nominate subspecies]
Kodada, Ján, Harman, Marcel and Selnekovič, Dávid. (2024). New records of the minute moss beetle Ochthebius (Enicocerus) colveranus Ferro, 1979 in Slovakia (Coleoptera, Hydraenidae). Check List 20(6): 1216-1221. https://doi.org/10.15560/20.5.1216
Kodada, Ján, Jäch, M. A., Selnekovič, D. and Ondrejková, K. (2022). New records of Hydraena schuleri Ganglbauer, 1901 from Slovakia (Coleoptera, Hydraenidae). Check List 18(3): 571-574. https://doi.org/10.15560/18.3.571
Májeková, J., Zaliberová, M. and Jehlík, V. (2013). Extinct species Ceratocephala testiculata (Crantz) Besser rediscovered in Slovakia after 44 years. Thaiszia 23(2): 141-145. [Abstract]
Olejková, Dagmar et al. (2022). Yellow vetchling (Lathyrus aphaca L.) and round-seeded vetchling (Lathyrus sphaericus Retz.) – two species rediscovered for the flora of Slovakia. Thaiszia - J. Bot. 32(1): 55-65.
https://doi.org/10.33542/TJB2022-1-04
South Africa
Spain
Angus, Robert B. et al. (2022). Rediscovery of Helophorus cincticollis Guillebeau (Coleoptera, Helophoridae) in Spain. In: Adrián Villastrigo, Andrés Millán, David Sánchez-Fernández, Javier Fresneda, Luis F. Valladares (eds.). Advances in aquatic and subterranean beetles research: a tribute to Ignacio Ribera. Suplementos del Boletín de la Asociación española de Entomología 4: 1-2.
Carballeira, Rafael. (2018). Rediscovery of Eunapius carteri (Bowerbank, 1863) in Western Europe. Turkish Journal of Zoology 42: 137-145.
Olariaga I., Paz-Bermúdez G., Calvo J., Etayo J., Prieto M. (2020). Rediscovery of the endangered lichen Pseudocyphellaria aurata (Lobariaceae, Ascomycota) in mainland Spain. Anales del Jardín Botánico de Madrid 77: e099. https://doi.org/10.3989/ajbm.2558
Ortiz, J. H., López, F. V., 2012: Redescubrimiento de Lavatera trimestris var. brachypoda. Revista de la Sociedad Gaditana de Historia Natural 6, 23–24.
Villegas, Francisco Jesús Cano and del la Haza, Elisa Zafra. (2007). Redescubrimiento de Apteromantis aptera (Fuente, 1894) (Dictyoptera, Mantidae) en la provincia de Córdoba (sur de España). Boletín Sociedad Entomológica Aragonesa 41: 449-450.
Author? (1994). Rediscovery of Pardel Lynx colony. Animals & Men 2: 6.
Catalonia
Sri Lanka
Darshana, T. W. R. (In Press, 2018). Rediscovery of the Spoon-billed Sandpiper Calidris pygmaea in Sri Lanka after 40 years. Indian BIRDS.
De Silva, H. S. D. R., Wijekoon, W. M. C. D., Wegiriya, H. C. E. and Bandara, K. V. S. N. (2023). Rediscovery of the endemic firefly, Harmatelia bilinia Walker (Coleoptera: Lampyridae: Ototretinae) from Ranmale Forest Reserve, Sri Lanka. Proceedings of 10th Ruhuna International Science & Technology Conference, University of Ruhuna, Matara, Sri Lanka.
Fernando, Denise R. M. et al. (2022). Assessment of plant diversity and foliar chemistry on the Sri Lankan ultramafics reveals inconsistencies in the metal hyperaccumulator trait. Ecological Research 37: 215-227. [Rinorea bengalensis]
Nanayakkara, R., N. Vishvanath and T. G. T. Kusuminda. (2012). Rediscovery of Pouch bearing sheath–tailed bat Saccolaimus saccolaimus Temminck (Chiroptera: Emballonuridae) from Sri Lanka after 75 years. Asian Journal of Conservation Biology 1(2): 134-137.
Ranasinghe, T. and Ghate, H. V. (2022). Rediscovery of Gardena melinarthrum Dohrn from Sri Lanka. Journal of Threatened Taxa 14(6): 21318-21320. htps://doi.org/10.11609/jot.8052.14.6.21318-21320
de Silva, Anslem, M. Madawala, A.M. Bauer & S. Karunarathna (2016). Rediscovery of the Frilled Tail Gecko Hemidactylus platyurus (Schneider, 1792) in Sri Lanka after more than 160 years. Journal of Threatened Taxa 8(14): 9663-9666.
de Silva, Anslem et al. (2017). An authentic record of Eutropis bibronii (Gray, 1838) (Reptilia: Scincidae) from Sri Lanka. Zootaxa 4329(2): 175-182. [Abstract]
Wijesundara, Siril, Ranasinghe, Subhani, Jayasinghe, Himesh, Gunawardena, Nadeeka, Fonseka, Gayani and Wijesooriya, Shivanthi. (2020). Angiosperms in Sri Lanka: Present status of Angiosperms in Sri Lanka, pp. 1-168. In: The National Red List 2020 - Conservation Status of the Flora of Sri Lanka. Sri Lanka: Biodiversity Secretariat, Ministry of Environment and the National Herbarium, Department of National Botanic Gardens. xviii + 254 pp.
Suriname
Jairam, Rawien. (2017). The Distribution of Taeniophallus nicagus (Colubridae) in Suriname with Some Information on Morphology. IRCF Reptiles & Amphibians 24(3): 177-179.
Sweden
Switzerland
Vuataz, Laurent, Stucki, P., Fauquet, L. and Bernard, R. (2021). Rediscovery of Stactobia eatoniella McLachlan, 1880 (Trichoptera, Hydroptilidae) in Switzerland after more than seventy years. Alpine Entomology 5: 55-60. https://doi.org/10.3897/alpento.5.67985
Syria (Syrian Arab Republic)
Omran, Yaman, Baakliny, Johnny and Davis, Mark A. (2024). Community Science Rediscovers the Syrian Spadefoot Toad, Pelobates syriacus, in War-Torn Regions of Syria. Herpetological Conservation and Biology 19(3): 386-394.
Taiwan
Chen, Ying Chun et al. (2017). Re-emergence of Lilium callosum Sieb. et Zucc. in Taiwan after a fire allows propagation and renews the possibility of conservation. Botanical Studies 58: 47. https://doi.org/10.1186/s40529-017-0202-x
Lee, C. F. and Tsou, M. H. (2020). Rediscovery of Philopona vibex (Erichson, 1834) (Insecta: Coleoptera: Chrysomelidae: Galerucinae: Alticini) in Taiwan, with notes on its biology. J. Taiwan Agric. Res. 69(4): 265-273.
Liu, Shih-Hui, Huang, Chau-Ching and Liao, Chun-Kuei. (2021). Rediscovery of an ‘Extinct’ species Scleria sumatrensis Retz. in Taiwan using both morphological and molecular authentications. Taiwania 66(3): 398-407. doi:10.6165/tai.2021.66.398
Mong-Huai Su, Sheng-Chieh Wu, Chang-Fu Hsieh, Sin-I Chen and Kuoh-Cheng Yang. (2003). Rediscovery of Quercus aliena Blume (Fagaceae) in Taiwan. Taiwania 48(2): 112-117.
Terada, Katsuyuki, Yeh, Lan-Wei and Wu, Wen-Jer. (2013). Notes on the Taiwanese Caraboidea (Coleoptera) III. Rediscovery of Eustra chinensis Bänninger (Paussinae: Ozaenini). Collection and Research 26: 31-36.
Yeh, Lan-Wei, Terada, Katsuyuki and Huang, Chung-Hsin. (2023). A Preliminary Faunistic Survey of Carabidae (Coleoptera) from Gaomei Wetlands
Wildlife Refuge, Taichung City, Taiwan. Taiwanese Journal of Entomological Studies 8(1): 1-7.
Tanzania
Mlingwa, C. O. F. (1996). A note on the rediscovery of the Sokoke Pipit Anthus sokokensis in the Pugu Hills, Tanzania. Bird Cons. Int. 6: 293-294.
Thailand
Kunya, K.; O.S.G. Pauwels; P. Sairum; A. Taokratok. (2011). Squamata, Gekkonidae, Ptychozoon trinotaterra Brown, 1999: Rediscovery in Thailand. Check List 7(6): 820-822.
Trinidad & Tobago
Muhammad, Saifudeen M. and Ramdeen, Bryan. (2025). Return of a ghost: Rediscovery of the Trinidad Mahogany Tarantula Tapinauchenius latipes in Trinidad, W.I. Living World (J. Trinidad and Tobago Field Naturalists’ Club) 2025: 53-55.
Turkey
Adıgüzel, N., Bani, B. & Mavi, Ö. (2011). Rediscovery of Dichoropetalum aureum (Umbelliferae) in South Anatolia (Turkey). Phyton 50(2): 221–230.
Akkaya, A. and Uğurtaş, İ. H. (2011). Rediscovery of Vipera ammodytes (Linnaeus, 1758) at Uludağ-Bursa, Turkey, after 62 years. Herpetozoa 24(3-4): 181-185.
Yakin, Batuhan Yaman et al. (2018). New records and rediscovery of some snakes from Gökçeada (Imbros), Turkey. Biharean Biologist 12(1): e171305.
Ukraine
Balashov, I., Yarotskaya, M., Filatova, J., Starichenko, I. and Kovalov, V. (2017). Terrestrial Molluscs of the Tsyr-Pripyat Area in Volyn (Northern Ukraine): the First Findings of the Threatened Snail Vertigo moulinsiana in Mainland Ukraine. Vestnik zoologii 51(2): 251-258.
United Arab Emirates
Ahmed, S., Alzaabi, R., Soorae, P., Shah, J., Al Hammadi, E., Pusey, R. & Al Dhaheri, S. 2016. Rediscovering the Arabian sand cat (Felis margarita harrisoni) after a gap of 10 years using camera traps in the Western Region of Abu Dhabi, United Arab Emirates. European Journal of Wildlife Research 62(5): 627-631.
Todorova, V. 2018. Eyes in the desert: Two rare species snapped in Abu Dhabi. The National, August 11th , 2018.
Uruguay
Brucher, H. (1976). Zur Wiederentdeckung von Solanum commersonii Dunal nach 200 Jahren am originalstandort in Uruguay. (Rediscovery of Solanum commersonii on the original habitat in Uruguay after 200 years). Angew. Bot. 50(3-4): 97-111.
Hassemer, G. and Marchesi, E. H. (2016). Rediscovery of Plantago commersoniana (Plantaginaceae), a rare and threatened species, after two centuries in Uruguay. Webbia 71(1): 121-126. [Abstract]
Natale, G. S.; R. Maneyro. (2008). Amphibia, Anura, Leiuperidae, Pleurodema bibroni: Rediscovery. Check List 4(1): 47-49.
USA
Baker, Claude D. et al. (1985). Rediscovery of the Spotted Darter, Etheostoma maculatum, in Indiana Waters: Blue River; Crawford, Harrison and Washington Counties; Ohio River Drainage, USA. Indiana Academy of Science 94: 603-605. [automatic download]
Boone DD, Fenwick GH, Hirst F. 1984 The rediscovery of Oxypolis canbyi on the Delmarva Peninsula. Bartonia, no.50. 21-22.
Calder, Dale R. et al. (In Press, 2014). Clava multicornis (Forsskål, 1775): rediscovery of a North Atlantic hydroid (Cnidaria, Hydrozoa, Anthoathecata) on the Pacific coast of North America. BioInvasions Records.
Heger, Thierry J. et al. (2011). Rediscovery of Nebela ansata (Amoebozoa: Arcellinida) in eastern North America: biogeographical implications. Journal of Biogeography 38: 1897-1906.
Morrill, Daniel P.; Rezac, Calvin R.; Adams, Ginny; Adams, S. R.; and Hrabik, Robert (2021). Rediscovery of the Pallid Shiner, Hybopsis amnis, in the Black River System of Arkansas and Missouri Including Notes on Ecology and Life History. Southeastern Fishes Council Proceedings 61(1): article 2.
Schmidt Silveira, Fernanda, Schneider, Angelo Alberto, de Moura Baptista, Luis Rios. (2019). The role of a local rediscovery in the evaluation of the conservation status of a plant species: Testing the hypothesis of the biodiversity knowledge gap. Journal for Nature Conservation 48: 91-98. [Abstract]
Skvarla, Michael J. and Fisher, J. Ray. (2022). Rediscovery of Polystoechotes punctata (Fabricius, 1793) (Neuroptera: Ithonidae) in Eastern North America. Proceedings of the Entomological Society of Washington 124(2): 332-345. https://doi.org/10.4289/0013-8797.124.2.332
Wood, Julia E. et al. (2021). Range Extension of Blackfin Darter and Tennessee Dace, and First Collection of Western Blacknose Dace from Locust Fork of the Black Warrior River in 80 Years. Southeastern Naturalist 20(1): N30-N36. [Abstract]
[author?]. (2002). Rediscovery of Labidesthes sicculus (Atherinidae) in Lake Texoma (Oklahoma-Texas). Southwestern Naturalist 47: 142-147.
[author?]. (2002). The rediscovery of the Mississippi silverside, Menidia audens, in the Pearl River drainage in Mississippi and Louisiana. [incomplete citation]
Alabama
England, J. Kevin et al. (2021). Rockefellera crossophylla (Pannariaceae) Rediscovered in Alabama. Opuscula Philolichenum 20: 1-6.
Keener, Brian R. and Davenport, L. J. (2013). Rediscovery of Callirhoe papaver (Malvaceae) in Alabama (U.S.A.). Journal of the Botanical Research Institute of Texas 7(2):835-839. [Abstract]
Alaska
Fryday, Alan M. and Tønsberg, Tor. (2015). Ameliella - a bryicolous lichen genus rediscovered in North America after 50 years. Evansia 32(4): 171-175. [Abstract]
Arizona
Bezy, Robert L., Sherbrooke, Wade C. and Lowe, Charles H. (1966). The rediscovery of Eleutherodactylus augusti in Arizona. Herpetologica 22(3): 221-225.
Boucher, P. F. (1988). Shorter notes: rediscovery of Gymnocarpium dryopteris in Arizona. Amer. Fern J. 78(2): 71.
Carnahan, Susan D. and Verrier, James T. (2022). Hosackia oblongifolia (Fabaceae) Rediscovered in Arizona after 111-year Absence. Desert Plants 37(2): 37-39.
Embrey, Teague. (2021). Rediscovery of Eriogonum heermannii var. argense in southeastern Arizona. Phytoneuron 1: 1-7.
Arkansas
Serviss, Brett E. and Peck, James H. (2013). Rediscovery of Persea borbonia var. borbonia (Lauraceae), Prosopis glandulosa var. glandulosa (Fabaceae), and Pinus palustris (Pinaceae) in Arkansas, with three new angiosperm species for Arkansas (U.S.A.). Journal of the Botanical Research Institute of Texas 7(2):841-845. [Abstract]
Skvarla, Michael J. and Fisher, J. Ray. (2022). Rediscovery of Polystoechotes punctata (Fabricius, 1793) (Neuroptera: Ithonidae) in Eastern North America. Proceedings of the Entomological Society of Washington 124(2): 332-345. https://doi.org/10.4289/0013-8797.124.2.332
California
Cowart, Dominique A. et al. (2018). Environmental DNA (eDNA) applications for the conservation of imperiled crayfish (Decapoda: Astacidea) through monitoring of invasive species barriers and relocated populations. Journal of Crustacean Biology. https://doi.org/10.1093/jcbiol/ruy007 [Pacifastacus fortis]
Howard, Alice. (1980). In search of the Yosemite onion. Fremontia 8(1): 15-18.
Kippenhan, Michael G., Knisley, C. Barry and Haines, R. Dennis. (2012). Rediscovery and status of Cylindera (s. str.) lunalonga (Schaupp, 1884) (Coleoptera: Carabidae: Cicindelinae) in the San Joaquin Valley of California with a comparison to a Sierra Nevada population. Insecta Mundi 0254: 1-27.
Rejmánek, Marcel. (2018). Vascular plant extinctions in California: A critical assessment. Diversity and Distributions 24(1): 129-136. [Pyrola chlorantha (Eldorado County, in 1964); Triteleia grandiflora (Mendocino County, 1993)]
Smith, James Payne, Jr., and Berg, Ken. (1988). Inventory of rare and endangered vascular plants of California. Special Publication No. 1 (4th edition). California Native Plant Society. Sacramento. xviii + 170 pp. [Diplacus pygmaeus]
Colorado
Bestgen, K. R., Fausch, K. D., & Riley, S. C. (1991). Rediscovery of a relict southern population of Lake Chub, Couesius plumbeus, in Colorado. The Southwestern Naturalist, 36(1), 125-127.
Connecticut
Anagnostakis, Sandra L., Welch, Kenneth M., Snow, J. Wendell, Scarborough, Kathy and Eichlin, Thomas D. (1994). The Rediscovery of the Clearwing Chestnut Moth, Synanthedon castaneae (Busck) (Lepidoptera: Sesiidae) in Connecticut. Journal of the New York Entomological Society 102(1): 111-112.
Wehr, John D., Steirer, Sarah E. and Sleith, Robin S. (2019). Rediscovery of the Freshwater Brown Alga Heribaudiella in Connecticut after 100 Years. Northeastern Naturalist, 26(2): 343-361. [Abstract]
Delaware
McAvoy, William A. (2021). Noteworthy plant collections from Delaware, Maryland, and Virginia. Phytoneuron 19: 1-8. [Carex lucorum and possibly Opuntia mesacantha mesacantha]
Lobelia boykinii [rediscovered after 100 years and 19 days]
Castanea dentata (American chestnut)
Florida
Alexander, T. R. (1974). Schizaea germanii rediscovered in Florida. Amer. Fern J. 64(1): 30.
Hribar, Lawrence J. and Cerminara, Carrie. (2021). Rediscovery of Aedes scapularis (Diptera: Culicidae) in the Florida Keys. Florida Entomologist 104(2): 145.
Ward, D. B. and Minno, M. C. (2002). Rediscovery of the endangered Okeechobee gourd (Cucurbita okeechobeensis) along the St. Johns river, Florida, where last reported by William Bartram in 1774. Castanea 67(2): 201-206. https://www.jstor.org/stable/4034196
Georgia
Beck, A. F. (1990). 1989 season summary. Zone 9 southeast. Georgia. News Lepid. Soc. 2: 28. [Chlosyne gorgone gorgone]
Bridges EL, Orzell SL. 1992 The rediscovery of Rhynchospora solitaria Harper (Cyperaceae) in Georgia. Phytologia 72. (5): 369-372.
Bruce JG. 1979 Vittaria lineata rediscovered in Georgia. Amer. Fern J. 69. (1): 29.
Stark, B. P., Harrison, A. B. and Nye, K. C. (2016). Records of an uncommon stonefly, Nemocapnia carolina Banks (Plecoptera: Capniidae), in South Carolina and nearby states. Illiesia 12(2): 10-14.
Wisniewski, J.M., Rankin, N.M., Weiler, D.A., Strickland, B.A. and H.C. Chandler. (2013). Use of occupancy modelling to assess the status and habitat relationships of freshwater mussels in the Lower Flint River, Georgia, USA. Walkerana 17(1): 24-40.
Hawaii
Abbott, J.C. & Petr, D. 1997. Rediscovery of Apterocyclus honoluluensis Waterhouse on Kauai (Coleoptera: lucanidae). Bishop Museum Occasional Papers 49(2): 71. [erroneous taxonomic assignation, i.e. not really A. honoluluensis]
Boynton, David and Wood, Kenneth R. (2007). Erinna newcombi Adams & Adams (Mollusca: Lymnaeidae): a Rediscovered Population in Hanakoa, Kaua'i, Hawai'i. Bishop Museum Occasional Papers 96: 52-54.
Hodel, Donald R. (2009). A new species of Pritchardia and the rediscovery of P. lowreyana on Oahu, Hawaii. Palms 53(4): 173-179.
Lorence, David H., Flynn, Timothy W. and Wagner, Warren L. (1995). Contributions to the flora of Hawai`i. III. New additions, range extensions, and rediscoveries of flowering plants. Bishop Museum Occasional Papers 41: 19-58. [Spermolepis hawaiiensis and Argemona glauca on Kauai]
Nyberg, Benjamin et al. (2023). Recent notable plant records and rediscoveries from Kaua‘i, Hawaiian Islands. Records of the Hawaii Biological Survey for 2022. Edited by Neal L. Evenhuis. Bishop Museum Occasional Papers 148: 163-168. [Gouania meyenii, Silene lanceolata]
Oppenheimer, H. L. (2008). New Hawaiian plant records for 2007. Bishop Mus. Occas. Pap. 100: 22-38. [Neraudia
melastomifolia; Trisetum inaequale (on Lanai); Pteris lidgatei (on Molokai)]
Polhemus, D.A., Oppenheimer, H.L., Starr, F. and Martz, K. (1999). Notable rediscoveries of Megalagrion species on Maui (odonata: Coenagrionidae). Bishop Museum Occasional Papers 59(2): 27-29. [M. pacificum, M. xanthomelas]
Preston, David and Imada, Clyde. (2014). Biological Survey on a 10-Acre Parcel and Adjacent Areas, Proposed Land Acquisition, Kahului, Maui, Hawaii. Final Report. Hawaii Biological Survey No. 2014-001. [rediscovery of Manduca blackburni in 1984 on Maui]
Strong, M. T. and Wagner, W. L. (1997). New and noteworthy Cyperaceae from the Hawaiian Islands. Bishop Mus. Occas. Pap. 48: 37-50. [Cyperus esculentus]
Wagner, W. L. and Herbst, D. R. (1995). Contributions to the flora of Hawai`i. IV. New records and name changes. Bishop Mus. Occas. Pap. 42: 13-27. [Spermolepis hawaiiensis on Lanai]
Wood, Kenneth R. (2006). New Plant Records and Rediscoveries within the Hawaiian Islands. Bishop Museum Occasional Papers 88: 15-19. [Diellia erecta alexandri]
Wood, Kenneth R. (2007). New plant records, rediscoveries, range extensions, and possible extinctions within the Hawaiian Islands. Bishop Mus. Occas. Pap. 96: 13-17. [Diplazium molokaiense]
Wood, Kenneth R. (2012). Posible Extinctions, Rediscoveries, and New Plant Records within the Hawaiian Islands. Records of the Hawaii Biological Survey for 2011. Edited by Neal L. Evenhuis & Lucius G. Eldredge. Bishop Museum Occasional Papers 113: 91-102. [Ctenitis squamigera, Lysimachia filifolia, Cyclosorus pendens]
Wood, Kenneth R., Burney, D. A., Allison, A. and Fisher, R. (2013). Emoia impar (Squamata, Scincidae): not extinct in the Hawaiian Islands. Oryx 47: 328.
Wood, Kenneth R. and Kirkpatrick, Megan. (2014). Rediscovery of Melicope quadrangularis (Rutaceae) and other notable plant records for the island of Kaua‘i, Hawai‘i. Records of the Hawaii Biological Survey for 2013. Edited by Neal L. Evenhuis. Bishop Museum Occasional Papers 115: 29-32. [Lysimachia filifolia]
Megalagrion xanthomelas (O'ahu in 1994)
Illinois
Bouseman, J. K. (1982). Note on the rediscovery of Xeromelecta interrupta (Cresson) in Illinois (Hymenoptera: Apoidea). Journal of the Kansas Entomological Society 55(2): 349-350.
Kluge, Mark and Garness, Kathleen Marie. (2022). Newly Documented Populations of Two State Endangered Carex Species in Lake County, Illinois. Erigenia 28: 74-76. [Carex intumescens (Cook County)]
Lynch, John T. (1965). Rediscovery of the Four-Toed Salamander, Hemidactglium scutatum, in Illinois: A Relict Population. Herpetologica 21(2): 152-153.
Pilla, N. J. and Anastasio, A., (2024). The Rediscovery of Eleocharis geniculata (L.) Roem. & Schult. (Cyperaceae) in Illinois with Taxonomic Notes. The Great Lakes Botanist 63(1-2): 4, 31-36. https://doi.org/10.3998/glbot.7050
Indiana
Alix MS, Scribailo RW. ("2001" 2002). Rediscovery of Wolffiella gladiata (Lemnaceae) in Indiana. Mich. Bot. 40. (1): 17-21.
Baker, Claude D., Forsyth, Bill J. and Wiles, Tom. (1985). Rediscovery of the Spotted Darter, Etheostoma maculatum, in Indiana Waters: Blue River; Crawford, Harrison and Washington Counties; Ohio River Drainage, USA. Indiana Academy of Science 94: 603-605.
Homoya, M. A., Aldrich, J. R. and Jacquart, E. M. (1989). The rediscovery of the globally endangered clover, Trifolium stoloniferum, in Indiana. Rhodora 91(866): 207-212.
Kansas
Angelo, R.T., and M.S. Cringan. (2003). Rediscovery of the black sandshell, Ligumia recta (Bamarck, 1819), in Kansas. Transactions of the Kansas Academy of Science 106(1-2): 111-113.
Gustafson, Grey T. et al. (2014). Rediscovery of Lutrochus laticeps Casey, 1893 (Coleoptera: Lutrochidae) and the Discovery of Dineutus productus Roberts, 1895 and Dineutus serrulatus analis Régimbart, 1882 (Coleoptera: Gyrinidae) in Kansas, USA, with Notes on Habitat Preference. The Coleopterists Bulletin 68(4): 714-718.
Kentucky
Baskin JM, Baskin CC. 1984 Rediscovery of the rare Kentucky endemic Solidago shortii T. & G. in Fleming and Nicholas counties. Trans. Kentucky Acad. Sci., 45(3-4): 159.
Eisenhour, David J.; Thomas, Matthew R.; Culp, J. Jacob; Compton, Michael C.; Brandt, Stephanie L.; and Pierce, Rodney. (2018). Updated Distributional Records of Selected Kentucky Fishes. Southeastern Fishes Council Proceedings 58: 1-19. [Ichthyomyzon gagei, Hemitremia flammea, Hybopsis amnis]
Louisiana
Doffitt, Christopher H. (2016). Rediscovery of Triosteum perfoliatum in Louisiana. Phytoneuron 56: 1-2.
Maine
Arsenault MP, Haines A. 2004 New England note. Rediscovery of Carex typhina (Cyperaceae) in Maine. Rhodora 106. (925): 52-54.
Daigle, J. J. (1988). Ophiogomphus anomalus Harvey rediscovered in Maine, United States (Anisoptera: Gomphidae). Notul. Odonatol. 3: 32-33.
Hellquist, C. B. (1981). The rediscovery of Juncus stygius in Maine. Rhodora 83(834): 321-322.
Maryland
Frye, Christopher, McAvoy, William and Thomas, Justin. (2022). Rediscovery of Dichanthelium ravenelii (Poaceae) in Maryland. Phytoneuron 56: 1-2.
McAvoy, William A. (2021). Noteworthy plant collections from Delaware, Maryland, and Virginia. Phytoneuron 19: 1-8. [Hexalectris spicata]
Minney, Coleman and Landsman, Andrew P. (In Press, 2017). Rediscovery of Ribes americanum (Grossulariaceae) in Maryland. Castanea.
Massachusetts
Angelo, R. (1979). Ledum groenlandicum rediscovered in Concord, Massachusetts. Rhodora 81: 285-286.
Davidson, Robert and Ryyken, Jessica. (2011). Rediscovery of Bembidion (Lymnaeum) nigropiceum (Marsham) (= puritanum Hayward) in Massachusetts, with remarks on biology and habitat (Coleoptera, Carabidae, Bembidiini). ZooKeys 147: 487-496.
Michigan
Taylor, S. M. (1990). The Allegheny plum of Michigan’s jack pine plains. Michigan Academician 22(4): 381-384. [Prunus alleghaniensis]
Mississippi
MacGown, J. A. and Hill, J. G. (2007). Rediscovery of the Florida harvester ant in mainland Mississippi. Mississippi Native Plants and Environmental Education Newletter 25: 1-4.
MacGown, J. A., Hill, J. G., Majure, L. C. and Seltzer, J. L. (2008). Rediscovery of Pogonomyrmex badius (Latreille) (Hymenoptera: Formicidae) in mainland Mississippi, U.S.A with an analysis of associated seeds and vegetation. Midsouth Entomology 1: 17-28.
Missouri
Brant, Alan E. (2019). Panic grasses of southeastern Missouri–one new and one rediscovery. Phytoneuron 14: 1-3.
Buback, Steven. (2021). Rediscovery of Echinacea angustifolia and Cirsium undulatum in the loess hills of Missouri, with confirmation of Solidago jejunifolia. Missouriensis 39: 1-6.
Wakeman, Ann and McKenzie, Paul M. (1996). Rediscovery of Oryzopsis racemosa (Poaceae) in Missouri. Missouriensis 16(2): 16-23.
New Hampshire
Brackley, F. E. (1979). Cypripedium reginae rediscovered in New Hampshire. Rhodora 81. (828): 579-580.
New Jersey
New Mexico
New York
Losey, John E. et al. (2014). Lady Beetles in New York: Insidious Invasions, Erstwhile Extirpations and Recent Rediscoveries. Northeastern Naturalist 21(2): 271-284. [Adalia bipunctata, Coccinella novemnotata]
Rosen, David J., Richardson, Alfred and King, Ken. (2016). Rediscovery of Eleocharis obtusetrigona (Cyperaceae) in Texas. Phytoneuron 45: 1-3.
Witherspoon, Caroline M. and Lendemer, James C. (2026). Noteworthy Records of Three Usnea Species from the New York Adirondacks. Evansia 42(4): 69-81. https://doi.org/10.1639/0747-9859-42.4.69 [Usnea ceratina, Usnea subfusca]
Damak-damak Tahun (Scolopia macrophylla)
Ohio
Cusick, A. W. (1988). Trifolium stoloniferum (Fabaceae) rediscovered in Ohio. Ohio J. Sci. 89(2): 6.
Cusick, A. W. (1989). Trifolium stoloniferum (Fabaceae) in Ohio: history, habitats, decline and rediscovery. Sida 13(4): 467-480.
Riley, B. P., Vincent, M. A. and Widrlechner, M. P. (2020). Ohio flora: Additions, noteworthy finds, and deletions. Phytoneuron 2020(2): 1-39. [Amelanchier interior, Anagallis minima, Carex formosa, Juncus subcaudatus, Rubus trivialis]
Curved tortella moss (Tortella inclinata)
Oregon
Kagan, Jimmy and Vrilakas, Susan. (1993). Extinct and Extirpated Plants from Oregon. Kalmiopsis [1993]: 12-16.
Siddall, Jean L., Chambers, Kenton L. and Wagner, David H. (1979). Rare, threatened and endangered vascular plants in Oregon — an interim report. Oregon Natural Area Preserves Advisory Committee, Division of State Lands. 109 pp. [Bensoniella oregona]
Pennsylvania
Buker W. 1980 Rediscovery of colic root in western Pennsylvania. Castanea 45. (2): 143-144. [Aletris farinosa]
Glon, Mael G. et al. (2018). Rediscovery of Cambarus diogenes (Devil Crayfish) in Pennsylvania. Northeastern Naturalist 25(3): 355-361.
Krochmal, Jimmy et al. (2022). The Rediscovery of the Spiny-cheek crayfish (Faxonius limosus) in the Middle and Upper Susquehanna River Basin in Pennsylvania. Proceedings of the West Virginia Academy of Science 94(1).
Tennessee
Ahlstedt, Steven A. et al. (2017). Freshwater mussels of the National Park Service Obed wild and scenic river, Tennessee. Malacological Review 45-46: 193-211. [Lampsilis virescens and Villosa perpurpurea]
Bates, V. and Lewis, P. (1984). Rediscovery of Stylisma humistrata (Convolvulaceae) in Tennessee. Rhodora 86(847): 393-394.
Floden, Aaron J. and Schilling, Edward E. (Accepted, 2017). Trautvetteria fonticalcarea (Ranunculaceae: Ranunculeae), a new tassel rue species endemic to Tennessee. Nordic Journal of Botany. DOI: 10.1111/njb.01738 [Abstract] [Rhynchospora capillacea]
Stout, Jeremy B. and Stout, Elliott S. (2022). Rediscovery and Site Ecology of Pennsylvania Catchfly (Silene caroliniana ssp. pensylvanica) at Watauga River Bluffs State Natural Area (Carter County, Tennessee). Castanea 87(2): 306-311.
Youssef, Nadeer N., Basham, Joshua P. and Oliver, Jason B. (2022). Rediscovery of Coccinella novemnotata Herbst (Coleoptera: Coccinellidae) in Tennessee, USA. The Coleopterists Bulletin 76(1): 82-83. https://doi.org/10.1649/0010-065X-76.1.82
Texas
Utah
Baumann, R. W. (2006). Rediscovery of the rare stonefly Utaperla sopladora at the type locality more than half a century after it was described. Western North American Naturalist, 66(2), 254-255.
Vermont
Congdon, J. W. (1875). Rare plants. Bulletin of the Torrey Botanical Club 6(9): 56.
Dobbin, Frank. (1907). Rediscovery of Podostemon ceratophyllus in Vermont. Rhodora 9(106): 220.
Pupko, Julia and McFarland, Kent. (2022). Vermont Lady Beetle Atlas: Progress Report. Vermont Center for Ecostudies. [Hippodamia convergens, Hyperaspis bigeminata, Brachiacantha quadripunctata, Hyperaspis octavia, Brachiacantha decempustulata, Mulsantina hudsonica]
Lythrum alatum (winged loosestrife): http://digital.vpr.net/post/rare-flower-thought-extinct-vermont-rediscovered#stream/0
Isotria medeoloides (Small whorled pogonia)
Empetrum atropurpureum (purple crowberry)
Wyoming
Marsh, P. C., Douglas, M. E., Minckley, W. L. and Timmons, R. J. (1991). Rediscovery of Colorado Squawfish, Ptychocheilus lucius (Cyprinidae), in Wyoming. Copeia 1991(4): 1091-1092.
Vietnam
Nguyen Quang Truong, Tran Thanh Tung, Hoang van Ngoc, Wolfgang Böhme, and Thomas Ziegler. (2008). Rediscovery and redescription of Ateuchosaurus chinensis Gray, 1845 (Squamata: Sauria: Scincidae) from northeastern Vietnam. Herpetology Notes 1: 17-21.
Orlov, Nikolai L., Ryabov, Sergei A., Nguyen, Tao Thien, and Nguyen, Truong Quang. (2010). Rediscovery and Redescription of Two Rare Snake Species: Oligodon lacroixi Angel et Bourret, 1933 and Maculophis bellus chapaensis (Bourret, 1934) [Squamata: Ophidia: Colubridae] from Fansipan Mountains, Northern Vietnam. Russian Journal of Herpetology 17(4): 310-322. [Abstract] [Oligodon lacroixi]
Trần, Hữu Đăng et al. (2018). Identification, sexual dimorphism and aspects of the natural history of Sapria himalayana (Rafflesiaceae) on Vietnam’s Lang Biang Plateau. Botanical Studies 59: 29.
Tu, V.T., S. Arai, F. Kikuchi, C.T. Hang, T.A. Tuan, G. Csorba & T. Gorfol (2019). Rediscovery of Van Hasselt’s Mouse-eared Bat Myots hasselti (Temminck, 1840) and its frst genetc data from Hanoi, northern Vietnam. Journal of Threatened Taxa 11(7): 13915-13919.
Wales
Blackstock, T. H. and Jones, R. A. (1997). Juncus capitatus Weigel (Juncaceae) rediscovered near its original locality in Anglesey (v.c. 52). Watsonia 21(3): 277-278.
Holloway, Graham J., Owen, Christian and Tordoff, George. (2024). Dermestes undulatus Brahm rediscovered in Wales – and its differentiation from Dermestes murinus Linnaeus (Coleoptera: Dermestidae). Entomologist's Monthly Magazine 160(4): 283-286. https://doi.org/10.31184/M00138908.1604.4239
Yugoslavia
Kuthan, Jan. (1980). Rediscovery of Stereum reflexulum on the Island of Lokrum (Yugoslavia) on another host. Acta Botanica Croatica 39(1): 143-144.
Zimbabwe
Boele, C. (1988). Brachystelma tavalla K.Schum rediscovered in Zimbabwe. Excelsa no.13. 47-49.
Appendix 1: Local rediscovery turned out to be new (sub)species
Watt, J. C. (1963). The rediscovery of a giant weta, Deinacrida heteracantha, on the North Island mainland. New Zealand Entomologist 3(2): 9-13.
Borya mirabilis: http://www.viridans.com/RAREPL/alwaysrare.htm
Appendix 2: Global rediscovery turned out to be new (sub)species
Ford, Bruce A. et al. (2017). Two Extraordinary New Pseudopetiolate Carex Species from Vietnam (sect. Hemiscaposae, Cyperaceae). Systematic Botany 42(3): 402-417. [Abstract]
Calystegia felix [originally thought to be C. sepium binghamiae]
Appendix 3: Country unknown
Silveira, Fernanda Schmidt et al. (In Press, 2018). The role of a local rediscovery in the evaluation of the conservation status of a plant species: Testing the hypothesis of the biodiversity knowledge gap. Journal for Nature Conservation. https://doi.org/10.1016/j.jnc.2018.10.004
Tamayo and D'Angelo. (1980). Hallazgo del Murcielago Colorado, Lasiurus borealis varius (Poeppig, 1835) en Magallanes y Cosideraciones Acerca de la Distribucion de los Lasiurus (Chiroptera: Vespertilionidae). Noticiario Mensual [1980]: 10. [Argentina or Chile]
Türkay, M. and Koukouras, A. (1988). The rediscovery of Monodaeus guinotae Forest, 1976 in the Aegean Sea. Senckenbergiana Biologica 68: 401-405.
Wasserlauf, Yohay et al. (2023). A telemetry study shows that an endangered nocturnal avian species roosts in extremely dry habitats to avoid predation. Scientific Reports 13: 11888. https://doi.org/10.1038/s41598-023-38981-2 [Egyptian Nightjar (Caprimulgus aegyptius)]
Appendix 4: Rediscovery turned out to be recolonisation
Manlick, Philip J. et al. (2018). Dynamic colonization history in a rediscovered Isle Royale carnivore. Scientific Reports 8: 12711.
Appendix 5: Local rediscovery rejected
Jumellea recurva on Mauritius:
Sevathian, J.C. (2011). The rediscovery of Jumellea recurva (Orchidaceae) in native forests of Bel Ombre, Mauritius. Phytotaxa: 29: 56–60.
Baider, Claudia et al. (2012). Jumellea recurva (Orchidaceae): not a rediscovery in Mauritius. Phytotaxa 40: 31-32. https://doi.org/10.11646/phytotaxa.40.1.5 [Abstract]
Amorphophallus staudtii from Ivory Coast:
Barthlott, W. and Bogner, J. (1981). Rediscovery of Amorphophallus staudtii (Engl.) N.E. Br. in the Tai National Park, Ivory Coast. Aroideana 4(4): 109-113.
Appendix 6: Rediscovery of introduced population
Costa, V. A. et al. (2022). Rediscovering Tetrastichus giffardianus (Hymenoptera: Eulophidae) 60 years after its introduction in Brazil. CArq. Inst. Biol. 72(4): 539-541. https://doi.org/10.1590/1808-1657v72p5392005